Filter
Associated Lab
- Ahrens Lab (5) Apply Ahrens Lab filter
- Aso Lab (1) Apply Aso Lab filter
- Baker Lab (2) Apply Baker Lab filter
- Betzig Lab (4) Apply Betzig Lab filter
- Bock Lab (2) Apply Bock Lab filter
- Cardona Lab (1) Apply Cardona Lab filter
- Cui Lab (2) Apply Cui Lab filter
- Dickson Lab (3) Apply Dickson Lab filter
- Druckmann Lab (1) Apply Druckmann Lab filter
- Dudman Lab (2) Apply Dudman Lab filter
- Eddy/Rivas Lab (2) Apply Eddy/Rivas Lab filter
- Egnor Lab (1) Apply Egnor Lab filter
- Fetter Lab (3) Apply Fetter Lab filter
- Fitzgerald Lab (1) Apply Fitzgerald Lab filter
- Gonen Lab (11) Apply Gonen Lab filter
- Grigorieff Lab (4) Apply Grigorieff Lab filter
- Harris Lab (3) Apply Harris Lab filter
- Heberlein Lab (6) Apply Heberlein Lab filter
- Hermundstad Lab (1) Apply Hermundstad Lab filter
- Hess Lab (2) Apply Hess Lab filter
- Jayaraman Lab (3) Apply Jayaraman Lab filter
- Ji Lab (1) Apply Ji Lab filter
- Johnson Lab (1) Apply Johnson Lab filter
- Karpova Lab (1) Apply Karpova Lab filter
- Keller Lab (10) Apply Keller Lab filter
- Lavis Lab (4) Apply Lavis Lab filter
- Leonardo Lab (3) Apply Leonardo Lab filter
- Lippincott-Schwartz Lab (11) Apply Lippincott-Schwartz Lab filter
- Looger Lab (10) Apply Looger Lab filter
- Magee Lab (3) Apply Magee Lab filter
- Menon Lab (3) Apply Menon Lab filter
- Pachitariu Lab (3) Apply Pachitariu Lab filter
- Pavlopoulos Lab (1) Apply Pavlopoulos Lab filter
- Reiser Lab (2) Apply Reiser Lab filter
- Riddiford Lab (5) Apply Riddiford Lab filter
- Romani Lab (1) Apply Romani Lab filter
- Rubin Lab (5) Apply Rubin Lab filter
- Satou Lab (2) Apply Satou Lab filter
- Scheffer Lab (3) Apply Scheffer Lab filter
- Schreiter Lab (7) Apply Schreiter Lab filter
- Sgro Lab (1) Apply Sgro Lab filter
- Singer Lab (9) Apply Singer Lab filter
- Spruston Lab (2) Apply Spruston Lab filter
- Stern Lab (6) Apply Stern Lab filter
- Sternson Lab (3) Apply Sternson Lab filter
- Svoboda Lab (10) Apply Svoboda Lab filter
- Tjian Lab (1) Apply Tjian Lab filter
- Truman Lab (3) Apply Truman Lab filter
- Turaga Lab (2) Apply Turaga Lab filter
- Turner Lab (2) Apply Turner Lab filter
- Wu Lab (3) Apply Wu Lab filter
- Zlatic Lab (2) Apply Zlatic Lab filter
Associated Project Team
Publication Date
- December 2013 (13) Apply December 2013 filter
- November 2013 (10) Apply November 2013 filter
- October 2013 (20) Apply October 2013 filter
- September 2013 (19) Apply September 2013 filter
- August 2013 (15) Apply August 2013 filter
- July 2013 (19) Apply July 2013 filter
- June 2013 (17) Apply June 2013 filter
- May 2013 (10) Apply May 2013 filter
- April 2013 (12) Apply April 2013 filter
- March 2013 (11) Apply March 2013 filter
- February 2013 (19) Apply February 2013 filter
- January 2013 (29) Apply January 2013 filter
- Remove 2013 filter 2013
Type of Publication
194 Publications
Showing 31-40 of 194 resultsThe second messenger cyclic AMP (cAMP) operates in discrete subcellular regions within which proteins that synthesize, break down or respond to the second messenger are precisely organized. A burgeoning knowledge of compartmentalized cAMP signaling is revealing how the local control of signaling enzyme activity impacts upon disease. The aim of this Cell Science at a Glance article and the accompanying poster is to highlight how misregulation of local cyclic AMP signaling can have pathophysiological consequences. We first introduce the core molecular machinery for cAMP signaling, which includes the cAMP-dependent protein kinase (PKA), and then consider the role of A-kinase anchoring proteins (AKAPs) in coordinating different cAMP-responsive proteins. The latter sections illustrate the emerging role of local cAMP signaling in four disease areas: cataracts, cancer, diabetes and cardiovascular diseases.
Fluorescent protein-based sensors for detecting neuronal activity have been developed largely based on non-neuronal screening systems. However, the dynamics of neuronal state variables (e.g., voltage, calcium, etc.) are typically very rapid compared to those of non-excitable cells. We developed an electrical stimulation and fluorescence imaging platform based on dissociated rat primary neuronal cultures. We describe its use in testing genetically-encoded calcium indicators (GECIs). Efficient neuronal GECI expression was achieved using lentiviruses containing a neuronal-selective gene promoter. Action potentials (APs) and thus neuronal calcium levels were quantitatively controlled by electrical field stimulation, and fluorescence images were recorded. Images were segmented to extract fluorescence signals corresponding to individual GECI-expressing neurons, which improved sensitivity over full-field measurements. We demonstrate the superiority of screening GECIs in neurons compared with solution measurements. Neuronal screening was useful for efficient identification of variants with both improved response kinetics and high signal amplitudes. This platform can be used to screen many types of sensors with cellular resolution under realistic conditions where neuronal state variables are in relevant ranges with respect to timing and amplitude.
Clusters of time series data may change location and memberships over time; in gene expression data, this occurs as groups of genes or samples respond differently to stimuli or experimental conditions at different times. In order to uncover this underlying temporal structure, we consider dynamic clusters with time-dependent parameters which split and merge over time, enabling cluster memberships to change. These interesting time-dependent structures are useful in understanding the development of organisms or complex organs, and could not be identified using traditional clustering methods. In cell cycle data, these time-dependent structure may provide links between genes and stages of the cell cycle, whilst in developmental data sets they may highlight key developmental transitions.
The manner in which different distributions of synaptic weights onto cortical neurons shape their spiking activity remains open. To characterize a homogeneous neuronal population, we use the master equation for generalized leaky integrate-and-fire neurons with shot-noise synapses. We develop fast semi-analytic numerical methods to solve this equation for either current or conductance synapses, with and without synaptic depression. We show that its solutions match simulations of equivalent neuronal networks better than those of the Fokker-Planck equation and we compute bounds on the network response to non-instantaneous synapses. We apply these methods to study different synaptic weight distributions in feed-forward networks. We characterize the synaptic amplitude distributions using a set of measures, called tail weight numbers, designed to quantify the preponderance of very strong synapses. Even if synaptic amplitude distributions are equated for both the total current and average synaptic weight, distributions with sparse but strong synapses produce higher responses for small inputs, leading to a larger operating range. Furthermore, despite their small number, such synapses enable the network to respond faster and with more stability in the face of external fluctuations.
Background: Segmenting electron microscopy (EM) images of cellular and subcellular processes in the nervous system is a key step in many bioimaging pipelines involving classification and labeling of ultrastructures. However, fully automated techniques to segment images are often susceptible to noise and heterogeneity in EM images (e.g. different histological preparations, different organisms, different brain regions, etc.). Supervised techniques to address this problem are often helpful but require large sets of training data, which are often difficult to obtain in practice, especially across many conditions. Results: We propose a new, principled unsupervised algorithm to segment EM images using a two-step approach: edge detection via salient watersheds following by robust region merging. We performed experiments to gather EM neuroimages of two organisms (mouse and fruit fly) using different histological preparations and generated manually curated ground-truth segmentations. We compared our algorithm against several state-of- the-art unsupervised segmentation algorithms and found superior performance using two standard measures of under-and over-segmentation error. Conclusions: Our algorithm is general and may be applicable to other large-scale segmentation problems for bioimages.
The suppression of tumorigenicity 2/IL-33 (ST2/IL-33) pathway has been implicated in several immune and inflammatory diseases. ST2 is produced as 2 isoforms. The membrane-bound isoform (ST2L) induces an immune response when bound to its ligand, IL-33. The other isoform is a soluble protein (sST2) that is thought to be a decoy receptor for IL-33 signaling. Elevated sST2 levels in serum are associated with an increased risk for cardiovascular disease. We investigated the determinants of sST2 plasma concentrations in 2,991 Framingham Offspring Cohort participants. While clinical and environmental factors explained some variation in sST2 levels, much of the variation in sST2 production was driven by genetic factors. In a genome-wide association study (GWAS), multiple SNPs within IL1RL1 (the gene encoding ST2) demonstrated associations with sST2 concentrations. Five missense variants of IL1RL1 correlated with higher sST2 levels in the GWAS and mapped to the intracellular domain of ST2, which is absent in sST2. In a cell culture model, IL1RL1 missense variants increased sST2 expression by inducing IL-33 expression and enhancing IL-33 responsiveness (via ST2L). Our data suggest that genetic variation in IL1RL1 can result in increased levels of sST2 and alter immune and inflammatory signaling through the ST2/IL-33 pathway.
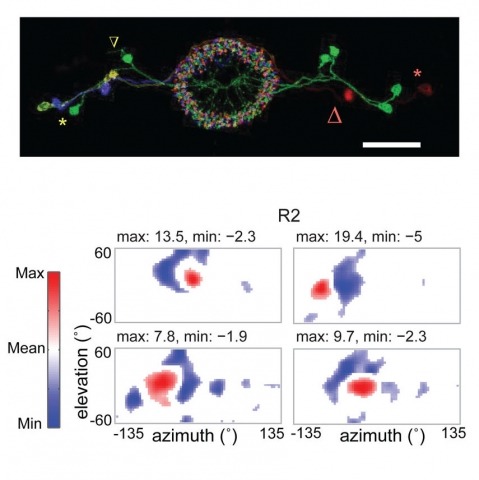
Many animals, including insects, are known to use visual landmarks to orient in their environment. In Drosophila melanogaster, behavioural genetics studies have identified a higher brain structure called the central complex as being required for the fly’s innate responses to vertical visual features and its short- and long-term memory for visual patterns. But whether and how neurons of the fly central complex represent visual features are unknown. Here we use two-photon calcium imaging in head-fixed walking and flying flies to probe visuomotor responses of ring neurons—a class of central complex neurons that have been implicated in landmark-driven spatial memory in walking flies and memory for visual patterns in tethered flying flies. We show that dendrites of ring neurons are visually responsive and arranged retinotopically. Ring neuron receptive fields comprise both excitatory and inhibitory subfields, resembling those of simple cells in the mammalian primary visual cortex. Ring neurons show strong and, in some cases, direction-selective orientation tuning, with a notable preference for vertically oriented features similar to those that evoke innate responses in flies. Visual responses were diminished during flight, but, in contrast with the hypothesized role of the central complex in the control of locomotion, not modulated during walking. Taken together, these results indicate that ring neurons represent behaviourally relevant visual features in the fly’s environment, enabling downstream central complex circuits to produce appropriate motor commands. More broadly, this study opens the door to mechanistic investigations of circuit computations underlying visually guided action selection in the Drosophila central complex.
Abstract We recently described a new form of neural integration and firing in a subset of interneurons, in which evoking hundreds of action potentials over tens of seconds to minutes produces a sudden barrage of action potentials lasting about a minute beyond the inciting stimulation. During this persistent firing, action potentials are generated in the distal axon and propagate retrogradely to the soma. To distinguish this from other forms of persistent firing, we refer to it here as ’retroaxonal barrage firing’, or ’barrage firing’ for short. Its induction is blocked by chemical inhibitors of gap junctions and curiously, stimulation of one interneuron in some cases triggers barrage firing in a nearby, unstimulated interneuron. Beyond these clues, the mechanisms of barrage firing are unknown. Here we report new results related to these mechanisms. Induction of barrage firing was blocked by lowering extracellular calcium, as long as normal action potential threshold was maintained, and it was inhibited by blocking L-type voltage-gated calcium channels. Despite its calcium dependence, barrage firing was not prevented by inhibiting chemical synaptic transmission. Furthermore, loading the stimulated/recorded interneuron with BAPTA did not block barrage firing, suggesting that the required calcium entry occurs in other cells. Finally, barrage firing was normal in mice with deletion of the primary gene for neuronal gap junctions (connexin36), suggesting that non-neuronal gap junctions may be involved. Together, these findings suggest that barrage firing is probably triggered by a multicellular mechanism involving calcium signalling and gap junctions, but operating independently of chemical synaptic transmission.
SUMMARY: Sequence database searches are an essential part of molecular biology, providing information about the function and evolutionary history of proteins, RNA molecules and DNA sequence elements. We present a tool for DNA/DNA sequence comparison that is built on the HMMER framework, which applies probabilistic inference methods based on hidden Markov models to the problem of homology search. This tool, called nhmmer, enables improved detection of remote DNA homologs, and has been used in combination with Dfam and RepeatMasker to improve annotation of transposable elements in the human genome. AVAILABILITY: nhmmer is a part of the new HMMER3.1 release. Source code and documentation can be downloaded from http://hmmer.org. HMMER3.1 is freely licensed under the GNU GPLv3 and should be portable to any POSIX-compliant operating system, including Linux and Mac OS/X. CONTACT: wheelert@janelia.hhmi.org.
We demonstrate a meaningful prospective power analysis for an (admittedly idealized) illustrative connectome inference task. Modeling neurons as vertices and synapses as edges in a simple random graph model, we optimize the trade-off between the number of (putative) edges identified and the accuracy of the edge identification procedure. We conclude that explicit analysis of the quantity/quality trade-off is imperative for optimal neuroscientific experimental design. In particular, identifying edges faster/more cheaply, but with more error, can yield superior inferential performance.