Filter
Associated Lab
- Aguilera Castrejon Lab (14) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (11) Apply Ahrens Lab filter
- Baker Lab (19) Apply Baker Lab filter
- Betzig Lab (12) Apply Betzig Lab filter
- Beyene Lab (5) Apply Beyene Lab filter
- Bock Lab (3) Apply Bock Lab filter
- Branson Lab (3) Apply Branson Lab filter
- Card Lab (6) Apply Card Lab filter
- Cardona Lab (19) Apply Cardona Lab filter
- Chklovskii Lab (3) Apply Chklovskii Lab filter
- Clapham Lab (1) Apply Clapham Lab filter
- Darshan Lab (4) Apply Darshan Lab filter
- Dennis Lab (1) Apply Dennis Lab filter
- Dickson Lab (14) Apply Dickson Lab filter
- Druckmann Lab (4) Apply Druckmann Lab filter
- Dudman Lab (12) Apply Dudman Lab filter
- Egnor Lab (7) Apply Egnor Lab filter
- Espinosa Medina Lab (4) Apply Espinosa Medina Lab filter
- Fetter Lab (10) Apply Fetter Lab filter
- Fitzgerald Lab (13) Apply Fitzgerald Lab filter
- Gonen Lab (32) Apply Gonen Lab filter
- Grigorieff Lab (28) Apply Grigorieff Lab filter
- Harris Lab (10) Apply Harris Lab filter
- Heberlein Lab (81) Apply Heberlein Lab filter
- Hermundstad Lab (4) Apply Hermundstad Lab filter
- Hess Lab (3) Apply Hess Lab filter
- Jayaraman Lab (4) Apply Jayaraman Lab filter
- Johnson Lab (5) Apply Johnson Lab filter
- Kainmueller Lab (19) Apply Kainmueller Lab filter
- Karpova Lab (1) Apply Karpova Lab filter
- Keleman Lab (5) Apply Keleman Lab filter
- Keller Lab (15) Apply Keller Lab filter
- Koay Lab (16) Apply Koay Lab filter
- Lavis Lab (12) Apply Lavis Lab filter
- Lee (Albert) Lab (5) Apply Lee (Albert) Lab filter
- Leonardo Lab (4) Apply Leonardo Lab filter
- Li Lab (24) Apply Li Lab filter
- Lippincott-Schwartz Lab (72) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (5) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (5) Apply Liu (Zhe) Lab filter
- Looger Lab (1) Apply Looger Lab filter
- Magee Lab (18) Apply Magee Lab filter
- Menon Lab (6) Apply Menon Lab filter
- Murphy Lab (7) Apply Murphy Lab filter
- O'Shea Lab (1) Apply O'Shea Lab filter
- Otopalik Lab (12) Apply Otopalik Lab filter
- Pachitariu Lab (12) Apply Pachitariu Lab filter
- Pastalkova Lab (13) Apply Pastalkova Lab filter
- Pavlopoulos Lab (12) Apply Pavlopoulos Lab filter
- Pedram Lab (11) Apply Pedram Lab filter
- Reiser Lab (6) Apply Reiser Lab filter
- Riddiford Lab (24) Apply Riddiford Lab filter
- Romani Lab (12) Apply Romani Lab filter
- Rubin Lab (38) Apply Rubin Lab filter
- Saalfeld Lab (17) Apply Saalfeld Lab filter
- Satou Lab (15) Apply Satou Lab filter
- Schreiter Lab (17) Apply Schreiter Lab filter
- Sgro Lab (20) Apply Sgro Lab filter
- Simpson Lab (5) Apply Simpson Lab filter
- Singer Lab (43) Apply Singer Lab filter
- Spruston Lab (36) Apply Spruston Lab filter
- Stern Lab (83) Apply Stern Lab filter
- Sternson Lab (7) Apply Sternson Lab filter
- Stringer Lab (3) Apply Stringer Lab filter
- Svoboda Lab (4) Apply Svoboda Lab filter
- Tebo Lab (24) Apply Tebo Lab filter
- Tillberg Lab (3) Apply Tillberg Lab filter
- Tjian Lab (47) Apply Tjian Lab filter
- Truman Lab (30) Apply Truman Lab filter
- Turaga Lab (12) Apply Turaga Lab filter
- Turner Lab (11) Apply Turner Lab filter
- Wang (Shaohe) Lab (19) Apply Wang (Shaohe) Lab filter
- Wu Lab (1) Apply Wu Lab filter
- Zlatic Lab (2) Apply Zlatic Lab filter
- Zuker Lab (20) Apply Zuker Lab filter
Associated Project Team
Publication Date
- 2023 (1) Apply 2023 filter
- 2022 (26) Apply 2022 filter
- 2021 (19) Apply 2021 filter
- 2020 (19) Apply 2020 filter
- 2019 (25) Apply 2019 filter
- 2018 (26) Apply 2018 filter
- 2017 (31) Apply 2017 filter
- 2016 (18) Apply 2016 filter
- 2015 (57) Apply 2015 filter
- 2014 (46) Apply 2014 filter
- 2013 (58) Apply 2013 filter
- 2012 (78) Apply 2012 filter
- 2011 (92) Apply 2011 filter
- 2010 (100) Apply 2010 filter
- 2009 (102) Apply 2009 filter
- 2008 (100) Apply 2008 filter
- 2007 (85) Apply 2007 filter
- 2006 (89) Apply 2006 filter
- 2005 (67) Apply 2005 filter
- 2004 (57) Apply 2004 filter
- 2003 (58) Apply 2003 filter
- 2002 (39) Apply 2002 filter
- 2001 (28) Apply 2001 filter
- 2000 (29) Apply 2000 filter
- 1999 (14) Apply 1999 filter
- 1998 (18) Apply 1998 filter
- 1997 (16) Apply 1997 filter
- 1996 (10) Apply 1996 filter
- 1995 (18) Apply 1995 filter
- 1994 (12) Apply 1994 filter
- 1993 (10) Apply 1993 filter
- 1992 (6) Apply 1992 filter
- 1991 (11) Apply 1991 filter
- 1990 (11) Apply 1990 filter
- 1989 (6) Apply 1989 filter
- 1988 (1) Apply 1988 filter
- 1987 (7) Apply 1987 filter
- 1986 (4) Apply 1986 filter
- 1985 (5) Apply 1985 filter
- 1984 (2) Apply 1984 filter
- 1983 (2) Apply 1983 filter
- 1982 (3) Apply 1982 filter
- 1981 (3) Apply 1981 filter
- 1980 (1) Apply 1980 filter
- 1979 (1) Apply 1979 filter
- 1976 (2) Apply 1976 filter
- 1973 (1) Apply 1973 filter
- 1970 (1) Apply 1970 filter
- 1967 (1) Apply 1967 filter
Type of Publication
- Remove Non-Janelia filter Non-Janelia
1416 Publications
Showing 1341-1350 of 1416 resultsIndividual carbocyanine dye molecules in a sub-monolayer spread have been imaged with near-field scanning optical microscopy. Molecules can be repeatedly detected and spatially localized (to approximately lambda/50 where lambda is the wavelength of light) with a sensitivity of at least 0.005 molecules/(Hz)(1/2) and the orientation of each molecular dipole can be determined. This information is exploited to map the electric field distribution in the near-field aperture with molecular spatial resolution.
Commentary: A paper of many firsts: the first single molecule microscopy; the first extended observations of single molecules under ambient conditions; the first localization of single molecules to near-molecular precision ( 15 nm), the first determination of the dipole axes of single fluorescent molecules; and the first near-molecular resolution optical microscopy, when a single fluorescent molecule was used to map the evanescent electric field components in the vicinity of a 100 nm diameter near-field aperture. Although eventually supplanted by simpler far-field methods, this paper ushered in the era of single molecule imaging and biophysics, and inspired the concept that would eventually lead to PALM. Even today, near-field single molecule detection lives on in the “zero mode waveguide” sequencing approach promoted by Pacific Biosciences.
Female sex determination in the germ line of Drosophila melanogaster is regulated by genes functioning in the soma as well as genes that function within the germ line. Genes known or suspected to be involved in germ-line sex determination in Drosophila melanogaster have been examined to determine if they are required upstream or downstream of Sex-lethal+, a known germ-line sex determination gene. Seven genes required for female-specific splicing of germ-line Sex-lethal+ pre-mRNA are identified. These results together with information about the tissues in which these genes function and whether they control sex determination and viability or just sex determination in the germ line have been used to deduce the genetic hierarchy regulating female germ-line sex determination. This hierarchy includes the somatic sex determination genes transformer+, transformer-2+ and doublesex+ (and by inference Sex-lethal+), which control a somatic signal required for female germ-line sex determination, and the germ-line ovarian tumor genes fused+, ovarian tumor+, ovo+, sans fille+, and Sex-lethal+, which are involved in either the reception or interpretation of this somatic sex determination signal. The fused+, ovarian tumor+, ovo+ and sans fille+ genes function upstream of Sex-lethal+ in the germ line.
Polarized angle-resolved Raman spectra of the Si-H stretching vibrations on stepped H-terminated Si(111) surfaces confirm the constrained orientation of the step dihydride derived from ab initio cluster calculations. They further show that the step normal modes involve little concerted motion of the step atoms, indicating that step relaxation reduces the steric interaction much further than predicted.
Near-field scanning optical microscopy (NSOM) has been used to generate high resolution flourescence images of cytoskeletal actin within fixed mouse fibroblast cells. Comparison with other microscopic methods indicates a transverse resolution well beyond that of confocal microscopy, and contrast far more revealing than in force microscopy. Effects unique to the near field are shown to be involved in the excitation of flourescence, yet the resulting images remain readily interpretable. As an initial demonstration of its utility, the technique is used to analyze the actin-based cytoskeletal structure between stress fibers and in cellular protrusions formed in the process of wound healing.
Commentary: The first superresolution fluorescence imaging of a biological system: the actin cytoskeleton in fixed, cultured fibroblast cells. This work strongly influenced me in two ways. First, calculations based on the signal-to-noise-ratio in images of single actin filaments in the paper suggested that single molecule imaging might be feasible. This was soon proven to be the case (see above). Second, the limitations of exogenous labeling for superresolution microscopy were revealed: samples which appeared correctly stained by conventional microscopy often exhibited sketchy, punctuate labeling of actin filaments as well as substantial non-specific background in the corresponding near field images. Indeed, it was the advent of GFP, with its promise of dense labeling and perfect specificity, that lured me back to superresolution microscopy when I first heard of it in 2003.
1. The voltage- and space-clamp errors associated with the use of a somatic electrode to measure current from dendritic synapses are evaluated using both equivalent-cylinder and morphologically realistic models of neuronal dendritic trees. 2. As a first step toward understanding the properties of synaptic current distortion under voltage-clamp conditions, the attenuation of step and sinusoidal voltage changes are evaluated in equivalent cylinder models. Demonstration of the frequency-dependent attenuation of voltage in the cable is then used as a framework for understanding the distortion of synaptic currents generated at sites remote from the somatic recording electrode and measured in the voltage-clamp recording configuration. 3. Increases in specific membrane resistivity (Rm) are shown to reduce steady-state voltage attenuation, while producing only minimal reduction in attenuation of transient voltage changes. Experimental manipulations that increase Rm therefore improve the accuracy of estimates of reversal potential for electrotonically remote synapses, but do not significantly reduce the attenuation of peak current. In addition, increases in Rm have the effect of slowing the kinetics of poorly clamped synaptic currents. 4. The effects of the magnitude of the synaptic conductance and its kinetics on the measured synaptic currents are also examined and discussed. The error in estimating parameters from measured synaptic currents is greatest for synapses with fast kinetics and large conductances. 5. A morphologically realistic model of a CA3 pyramidal neuron is used to demonstrate the generality of the conclusions derived from equivalent cylinder models. The realistic model is also used to fit synaptic currents generated by stimulation of mossy fiber (MF) and commissural/associational (C/A) inputs to CA3 neurons and to estimate the amount of distortion of these measured currents. 6. Anatomic data from the CA3 pyramidal neuron model are used to construct a simplified two-cylinder CA3 model. This model is used to estimate the electrotonic distances of MF synapses (which are located proximal to the soma) and perforant path (PP) synapses (which are located at the distal ends of the apical dendrites) and the distortion of synaptic current parameters measured for these synapses. 7. Results from the equivalent-cylinder models, the morphological CA3 model, and the simplified CA3 model all indicate that the amount of distortion of synaptic currents increases steeply as a function of distance from the soma. MF synapses close to the soma are likely to be subject only to small space-clamp errors, whereas MF synapses farther from the soma are likely to be substantially attenuated.(ABSTRACT TRUNCATED AT 400 WORDS)
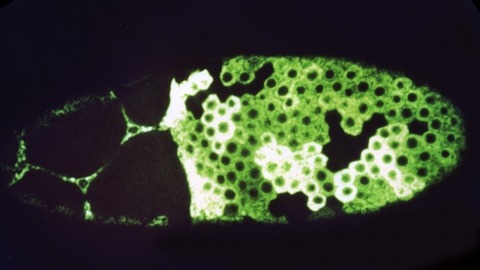
We have constructed a series of strains to facilitate the generation and analysis of clones of genetically distinct cells in developing and adult tissues of Drosophila. Each of these strains carries an FRT element, the target for the yeast FLP recombinase, near the base of a major chromosome arm, as well as a gratuitous cell-autonomous marker. Novel markers that carry epitope tags and that are localized to either the cell nucleus or cell membrane have been generated. As a demonstration of how these strains can be used to study a particular gene, we have analyzed the developmental role of the Drosophila EGF receptor homolog. Moreover, we have shown that these strains can be utilized to identify new mutations in mosaic animals in an efficient and unbiased way, thereby providing an unprecedented opportunity to perform systematic genetic screens for mutations affecting many biological processes.
Human mitochondrial transcription factor A is a 25-kDa protein that binds immediately upstream of the two major mitochondrial promoters, thereby leading to correct and efficient initiation of transcription. Although the nature of yeast mitochondrial promoters is significantly different from that of human promoters, a potential functional homolog of the human transcriptional activator protein has been previously identified in yeast mitochondria. The importance of the yeast protein in yeast mitochondrial DNA function has been shown by inactivation of its nuclear gene (ABF2) in Saccharomyces cerevisiae cells resulting in loss of mitochondrial DNA. We report here that the nuclear gene for human mitochondrial transcription factor A can be stably expressed in yeast cells devoid of the yeast homolog protein. The human protein is imported efficiently into yeast mitochondria, is processed correctly, and rescues the loss-of-mitochondrial DNA phenotype in a yeast abf2 strain, thus functionally substituting for the yeast protein. Both human and yeast proteins affect yeast mitochondrial transcription initiation in vitro, suggesting that the two proteins may have a common role in this fundamental process.
A new and conceptually simple data structure, called a suffix array, for on-line string searches is introduced in this paper. Constructing and querying suffix arrays is reduced to a sort and search paradigm that employs novel algorithms. The main advantage of suffix arrays over suffix trees is that, in practice, they use three to five times less space. From a complexity standpoint, suffix arrays permit on-line string searches of the type, ‘‘Is W a substring of A?’’ to be answered in time O(P + log N), where P is the length of W and N is the length of A, which is competitive with (and in some cases slightly better than) suffix trees. The only drawback is that in those instances where the underlying alphabet is finite and small, suffix trees can be constructed in O(N) time in the worst case, versus O(N log N) time for suffix arrays. However, we give an augmented algorithm that, regardless of the alphabet size, constructs suffix arrays in O(N) expected time, albeit with lesser space efficiency. We believe that suffix arrays will prove to be better in practice than suffix trees for many applications.
Recent advances in probe design have led to enhanced resolution (currently as significant as 12 nm) in optical microscopes based on near-field imaging. We demonstrate that the polarization of emitted and detected light in such microscopes can be manipulated sensitively to generate contrast. We show that the contrast on certain patterns is consistent with a simple interpretation of the requisite boundary conditions, whereas in other cases a more complicated interaction between the probe and the sample is involved. Finally application of the technique to near-filed magneto-optic imaging is demonstrated.
The near-field optical interaction between a sharp probe and a sample of interest can be exploited to image, spectroscopically probe, or modify surfaces at a resolution (down to approximately 12 nm) inaccessible by traditional far-field techniques. Many of the attractive features of conventional optics are retained, including noninvasiveness, reliability, and low cost. In addition, most optical contrast mechanisms can be extended to the near-field regime, resulting in a technique of considerable versatility. This versatility is demonstrated by several examples, such as the imaging of nanometric-scale features in mammalian tissue sections and the creation of ultrasmall, magneto-optic domains having implications for highdensity data storage. Although the technique may find uses in many diverse fields, two of the most exciting possibilities are localized optical spectroscopy of semiconductors and the fluorescence imaging of living cells.
Commentary: An overview of our work in near-field optics at the time, after our invention of the adiabatically tapered fiber probe and shear force feedback (see below) led to the first practical near-field scanning optical microscope. In this work, superresolution imaging via absorption, reflectivity, fluorescence, spectroscopy, polarization, and refractive index contrast were all demonstrated. Unlike all far-field superresolution fluorescence methods that were to appear a decade later, near-field microscopy remains the only superresolution technique capable of taking advantage of the full panoply of optical contrast mechanisms.