Filter
Associated Lab
- Betzig Lab (8) Apply Betzig Lab filter
- Eddy/Rivas Lab (2) Apply Eddy/Rivas Lab filter
- Fetter Lab (1) Apply Fetter Lab filter
- Hess Lab (2) Apply Hess Lab filter
- Ji Lab (4) Apply Ji Lab filter
- Looger Lab (3) Apply Looger Lab filter
- Magee Lab (4) Apply Magee Lab filter
- Riddiford Lab (1) Apply Riddiford Lab filter
- Rubin Lab (2) Apply Rubin Lab filter
- Schreiter Lab (1) Apply Schreiter Lab filter
- Shroff Lab (6) Apply Shroff Lab filter
- Spruston Lab (3) Apply Spruston Lab filter
- Sternson Lab (1) Apply Sternson Lab filter
- Svoboda Lab (6) Apply Svoboda Lab filter
- Truman Lab (1) Apply Truman Lab filter
Publication Date
- Remove 2007-12-31 19:00 – 2008-12-31 19:00 filter 2007-12-31 19:00 – 2008-12-31 19:00
- December 2008 (8) Apply December 2008 filter
- November 2008 (1) Apply November 2008 filter
- October 2008 (1) Apply October 2008 filter
- September 2008 (2) Apply September 2008 filter
- August 2008 (3) Apply August 2008 filter
- July 2008 (4) Apply July 2008 filter
- June 2008 (1) Apply June 2008 filter
- May 2008 (3) Apply May 2008 filter
- April 2008 (1) Apply April 2008 filter
- March 2008 (5) Apply March 2008 filter
- February 2008 (5) Apply February 2008 filter
- January 2008 (6) Apply January 2008 filter
40 Janelia Publications
Showing 31-40 of 40 resultsWe performed patch-clamp recordings from morphologically identified and anatomically mapped pyramidal neurons of the ventral hippocampus to test the hypothesis that bursting neurons are distributed on a gradient from the CA2/CA1 border (proximal) through the subiculum (distal), with more bursting observed at distal locations. We find that the well-defined morphological boundaries between the hippocampal subregions CA1 and subiculum do not correspond to abrupt changes in electrophysiological properties. Rather, we observed that the percentage of bursting neurons is linearly correlated with position in the proximal-distal axis across the CA1 and the subiculum, the percentages of bursting neurons being 10% near the CA1-CA2 border, 24% at the CA1-subiculum border, and higher than 50% in the distal subiculum. The distribution of bursting neurons was paralleled by a gradient in afterdepolarization (ADP) amplitude. We also tested the hypothesis that there was an association between bursting and two previously described morphologically distinct groups of pyramidal neurons (twin and single apical dendrites) in the CA1 region. We found no difference in output mode between single and twin apical dendrite morphologies, which was consistent with the observation that the two morphologies were equally distributed across the transverse axis of the CA1 region. Taken together with the known organization of connections from CA3 to CA1 and CA1 to subiculum, our results indicate that bursting neurons are most likely to be connected to regular spiking neurons and vice versa.
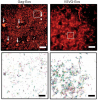
We combined photoactivated localization microscopy (PALM) with live-cell single-particle tracking to create a new method termed sptPALM. We created spatially resolved maps of single-molecule motions by imaging the membrane proteins Gag and VSVG, and obtained several orders of magnitude more trajectories per cell than traditional single-particle tracking enables. By probing distinct subsets of molecules, sptPALM can provide insight into the origins of spatial and temporal heterogeneities in membranes.
Commentary: As a stepping stone to true live cell PALM (see above), our collaborator Jennifer Lippincott-Schwartz suggested using the sparse photoactivation principle of PALM to track the nanoscale motion of thousands of individual molecules within a single living cell. Termed single particle tracking PALM (sptPALM), Jennifer’s postdocs Suliana Manley and Jen Gillette used the method in our PALM rig to create spatially resolved maps of diffusion rates in the plasma membrane of live cells. sptPALM is a powerful tool to study the active cytoskeletal or passive diffusional transport of individual molecules with far more measurements per cell than is possible without sparse photoactivation.
Pulsed lasers are key elements in nonlinear bioimaging techniques such as two-photon fluorescence excitation (TPE) microscopy. Typically, however, only a percent or less of the laser power available can be delivered to the sample before photoinduced damage becomes excessive. Here we describe a passive pulse splitter that converts each laser pulse into a fixed number of sub-pulses of equal energy. We applied the splitter to TPE imaging of fixed mouse brain slices labeled with GFP and show that, in different power regimes, the splitter can be used either to increase the signal rate more than 100-fold or to reduce the rate of photobleaching by over fourfold. In living specimens, the gains were even greater: a ninefold reduction in photobleaching during in vivo imaging of Caenorhabditis elegans larvae, and a six- to 20-fold decrease in the rate of photodamage during calcium imaging of rat hippocampal brain slices.

Pulsed lasers are key elements in nonlinear bioimaging techniques such as two-photon fluorescence excitation (TPE) microscopy. Typically, however, only a percent or less of the laser power available can be delivered to the sample before photoinduced damage becomes excessive. Here we describe a passive pulse splitter that converts each laser pulse into a fixed number of sub-pulses of equal energy. We applied the splitter to TPE imaging of fixed mouse brain slices labeled with GFP and show that, in different power regimes, the splitter can be used either to increase the signal rate more than 100-fold or to reduce the rate of photobleaching by over fourfold. In living specimens, the gains were even greater: a ninefold reduction in photobleaching during in vivo imaging of Caenorhabditis elegans larvae, and a six- to 20-fold decrease in the rate of photodamage during calcium imaging of rat hippocampal brain slices.
Commentary: Na Ji came to me early in her postdoc with an idea to reduce photodamage in nonlinear microscopy by splitting the pulses from an ultrafast laser into multiple subpulses of reduced energy. In six weeks, we constructed a prototype pulse splitter and obtained initial results confirming the validity of her vision. Further experiments with Jeff Magee demonstrated that the splitter could be used to increase imaging speed or reduce photodamage in two photon microscopy by one to two orders of magnitude. This project is a great example of how quickly one can react and exploit new ideas in the Janelia environment.
MOTIVATION: Caenorhabditis elegans, a roundworm found in soil, is a widely studied model organism with about 1000 cells in the adult. Producing high-resolution fluorescence images of C.elegans to reveal biological insights is becoming routine, motivating the development of advanced computational tools for analyzing the resulting image stacks. For example, worm bodies usually curve significantly in images. Thus one must ’straighten’ the worms if they are to be compared under a canonical coordinate system. RESULTS: We develop a worm straightening algorithm (WSA) that restacks cutting planes orthogonal to a ’backbone’ that models the anterior-posterior axis of the worm. We formulate the backbone as a parametric cubic spline defined by a series of control points. We develop two methods for automatically determining the locations of the control points. Our experimental methods show that our approaches effectively straighten both 2D and 3D worm images.
Electrical microstimulation can establish causal links between the activity of groups of neurons and perceptual and cognitive functions. However, the number and identities of neurons microstimulated, as well as the number of action potentials evoked, are difficult to ascertain. To address these issues we introduced the light-gated algal channel channelrhodopsin-2 (ChR2) specifically into a small fraction of layer 2/3 neurons of the mouse primary somatosensory cortex. ChR2 photostimulation in vivo reliably generated stimulus-locked action potentials at frequencies up to 50 Hz. Here we show that naive mice readily learned to detect brief trains of action potentials (five light pulses, 1 ms, 20 Hz). After training, mice could detect a photostimulus firing a single action potential in approximately 300 neurons. Even fewer neurons (approximately 60) were required for longer stimuli (five action potentials, 250 ms). Our results show that perceptual decisions and learning can be driven by extremely brief epochs of cortical activity in a sparse subset of supragranular cortical pyramidal neurons.
We present an algorithm for automatic segmentation of the human pelvic bones from CT datasets that is based on the application of a statistical shape model. The proposed method is divided into three steps: 1) The averaged shape of the pelvis model is initially placed within the CT data using the Generalized Hough Transform, 2) the statistical shape model is then adapted to the image data by a transformation and variation of its shape modes, and 3) a final free-form deformation step based on optimal graph searching is applied to overcome the restrictive character of the statistical shape representation. We thoroughly evaluated the method on 50 manually segmented CT datasets by performing a leave-one-out study. The Generalized Hough Transform proved to be a reliable method for an automatic initial placement of the shape model within the CT data. Compared to the manual gold standard segmentations, our automatic segmentation approach produced an average surface distance of 1.2 ± 0.3mm after the adaptation of the statistical shape model, which could be reduced to 0.7±0.3mm using a final free-form deformation step. Together with an average segmentation time of less than 5 minutes, the results of our study indicate that our method meets the requirements of clinical routine.
Neurons and glia are functionally organized into circuits and higher-order structures via synaptic connectivity, well-orchestrated molecular signaling, and activity-dependent refinement. Such organization allows the precise information processing required for complex behaviors. Disruption of nervous systems by genetic deficiency or events such as trauma or environmental exposure may produce a diseased state in which certain aspects of inter-neuron signaling are impaired. Optical imaging techniques allow the direct visualization of individual neurons in a circuit environment. Imaging probes specific for given biomolecules may help elucidate their contribution to proper circuit function. Genetically encoded sensors can visualize trafficking of particular molecules in defined neuronal populations, non-invasively in intact brain or reduced preparations. Sensor analysis in healthy and diseased brains may reveal important differences and shed light on the development and progression of nervous system disorders. We review the field of genetically encoded sensors for molecules and cellular events, and their potential applicability to the study of nervous system disease.
Pfam is a comprehensive collection of protein domains and families, represented as multiple sequence alignments and as profile hidden Markov models. The current release of Pfam (22.0) contains 9318 protein families. Pfam is now based not only on the UniProtKB sequence database, but also on NCBI GenPept and on sequences from selected metagenomics projects. Pfam is available on the web from the consortium members using a new, consistent and improved website design in the UK (http://pfam.sanger.ac.uk/), the USA (http://pfam.janelia.org/) and Sweden (http://pfam.sbc.su.se/), as well as from mirror sites in France (http://pfam.jouy.inra.fr/) and South Korea (http://pfam.ccbb.re.kr/).
The conditional expression of hairpin constructs in Drosophila melanogaster has emerged in recent years as a method of choice in functional genomic studies. To date, upstream activating site-driven RNA interference constructs have been inserted into the genome randomly using P-element-mediated transformation, which can result in false negatives due to variable expression. To avoid this problem, we have developed a transgenic RNA interference vector based on the phiC31 site-specific integration method.