Filter
Associated Lab
- Ahrens Lab (41) Apply Ahrens Lab filter
- Aso Lab (39) Apply Aso Lab filter
- Baker Lab (19) Apply Baker Lab filter
- Betzig Lab (98) Apply Betzig Lab filter
- Beyene Lab (4) Apply Beyene Lab filter
- Bock Lab (14) Apply Bock Lab filter
- Branson Lab (45) Apply Branson Lab filter
- Card Lab (32) Apply Card Lab filter
- Cardona Lab (44) Apply Cardona Lab filter
- Chklovskii Lab (10) Apply Chklovskii Lab filter
- Clapham Lab (10) Apply Clapham Lab filter
- Cui Lab (19) Apply Cui Lab filter
- Darshan Lab (8) Apply Darshan Lab filter
- Dickson Lab (32) Apply Dickson Lab filter
- Druckmann Lab (21) Apply Druckmann Lab filter
- Dudman Lab (34) Apply Dudman Lab filter
- Eddy/Rivas Lab (30) Apply Eddy/Rivas Lab filter
- Egnor Lab (4) Apply Egnor Lab filter
- Espinosa Medina Lab (12) Apply Espinosa Medina Lab filter
- Feliciano Lab (6) Apply Feliciano Lab filter
- Fetter Lab (31) Apply Fetter Lab filter
- Fitzgerald Lab (14) Apply Fitzgerald Lab filter
- Freeman Lab (15) Apply Freeman Lab filter
- Funke Lab (33) Apply Funke Lab filter
- Gonen Lab (59) Apply Gonen Lab filter
- Grigorieff Lab (34) Apply Grigorieff Lab filter
- Harris Lab (48) Apply Harris Lab filter
- Heberlein Lab (13) Apply Heberlein Lab filter
- Hermundstad Lab (17) Apply Hermundstad Lab filter
- Hess Lab (65) Apply Hess Lab filter
- Ilanges Lab (1) Apply Ilanges Lab filter
- Jayaraman Lab (39) Apply Jayaraman Lab filter
- Ji Lab (32) Apply Ji Lab filter
- Johnson Lab (1) Apply Johnson Lab filter
- Karpova Lab (13) Apply Karpova Lab filter
- Keleman Lab (8) Apply Keleman Lab filter
- Keller Lab (60) Apply Keller Lab filter
- Lavis Lab (120) Apply Lavis Lab filter
- Lee (Albert) Lab (29) Apply Lee (Albert) Lab filter
- Leonardo Lab (19) Apply Leonardo Lab filter
- Li Lab (1) Apply Li Lab filter
- Lippincott-Schwartz Lab (84) Apply Lippincott-Schwartz Lab filter
- Liu (Zhe) Lab (51) Apply Liu (Zhe) Lab filter
- Looger Lab (136) Apply Looger Lab filter
- Magee Lab (31) Apply Magee Lab filter
- Menon Lab (12) Apply Menon Lab filter
- Murphy Lab (6) Apply Murphy Lab filter
- O'Shea Lab (3) Apply O'Shea Lab filter
- Otopalik Lab (1) Apply Otopalik Lab filter
- Pachitariu Lab (28) Apply Pachitariu Lab filter
- Pastalkova Lab (5) Apply Pastalkova Lab filter
- Pavlopoulos Lab (7) Apply Pavlopoulos Lab filter
- Pedram Lab (1) Apply Pedram Lab filter
- Podgorski Lab (16) Apply Podgorski Lab filter
- Reiser Lab (43) Apply Reiser Lab filter
- Riddiford Lab (20) Apply Riddiford Lab filter
- Romani Lab (28) Apply Romani Lab filter
- Rubin Lab (101) Apply Rubin Lab filter
- Saalfeld Lab (41) Apply Saalfeld Lab filter
- Satou Lab (1) Apply Satou Lab filter
- Scheffer Lab (36) Apply Scheffer Lab filter
- Schreiter Lab (44) Apply Schreiter Lab filter
- Shroff Lab (20) Apply Shroff Lab filter
- Simpson Lab (18) Apply Simpson Lab filter
- Singer Lab (37) Apply Singer Lab filter
- Spruston Lab (55) Apply Spruston Lab filter
- Stern Lab (67) Apply Stern Lab filter
- Sternson Lab (47) Apply Sternson Lab filter
- Stringer Lab (23) Apply Stringer Lab filter
- Svoboda Lab (131) Apply Svoboda Lab filter
- Tebo Lab (7) Apply Tebo Lab filter
- Tervo Lab (9) Apply Tervo Lab filter
- Tillberg Lab (12) Apply Tillberg Lab filter
- Tjian Lab (17) Apply Tjian Lab filter
- Truman Lab (58) Apply Truman Lab filter
- Turaga Lab (34) Apply Turaga Lab filter
- Turner Lab (24) Apply Turner Lab filter
- Vale Lab (6) Apply Vale Lab filter
- Voigts Lab (1) Apply Voigts Lab filter
- Wang (Meng) Lab (7) Apply Wang (Meng) Lab filter
- Wang (Shaohe) Lab (1) Apply Wang (Shaohe) Lab filter
- Wu Lab (8) Apply Wu Lab filter
- Zlatic Lab (26) Apply Zlatic Lab filter
- Zuker Lab (5) Apply Zuker Lab filter
Associated Project Team
- CellMap (1) Apply CellMap filter
- COSEM (3) Apply COSEM filter
- Fly Descending Interneuron (10) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (14) Apply Fly Functional Connectome filter
- Fly Olympiad (5) Apply Fly Olympiad filter
- FlyEM (49) Apply FlyEM filter
- FlyLight (46) Apply FlyLight filter
- GENIE (40) Apply GENIE filter
- Integrative Imaging (1) Apply Integrative Imaging filter
- Larval Olympiad (2) Apply Larval Olympiad filter
- MouseLight (16) Apply MouseLight filter
- NeuroSeq (1) Apply NeuroSeq filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (22) Apply Tool Translation Team (T3) filter
- Transcription Imaging (45) Apply Transcription Imaging filter
Associated Support Team
- Anatomy and Histology (18) Apply Anatomy and Histology filter
- Cryo-Electron Microscopy (33) Apply Cryo-Electron Microscopy filter
- Electron Microscopy (10) Apply Electron Microscopy filter
- Fly Facility (39) Apply Fly Facility filter
- Gene Targeting and Transgenics (10) Apply Gene Targeting and Transgenics filter
- Integrative Imaging (10) Apply Integrative Imaging filter
- Janelia Experimental Technology (35) Apply Janelia Experimental Technology filter
- Management Team (1) Apply Management Team filter
- Molecular Genomics (15) Apply Molecular Genomics filter
- Primary & iPS Cell Culture (13) Apply Primary & iPS Cell Culture filter
- Project Technical Resources (31) Apply Project Technical Resources filter
- Quantitative Genomics (18) Apply Quantitative Genomics filter
- Scientific Computing Software (56) Apply Scientific Computing Software filter
- Scientific Computing Systems (6) Apply Scientific Computing Systems filter
- Viral Tools (14) Apply Viral Tools filter
- Vivarium (6) Apply Vivarium filter
Publication Date
- 2024 (87) Apply 2024 filter
- 2023 (177) Apply 2023 filter
- 2022 (166) Apply 2022 filter
- 2021 (174) Apply 2021 filter
- 2020 (178) Apply 2020 filter
- 2019 (177) Apply 2019 filter
- 2018 (206) Apply 2018 filter
- 2017 (186) Apply 2017 filter
- 2016 (191) Apply 2016 filter
- 2015 (195) Apply 2015 filter
- 2014 (190) Apply 2014 filter
- 2013 (136) Apply 2013 filter
- 2012 (112) Apply 2012 filter
- 2011 (98) Apply 2011 filter
- 2010 (61) Apply 2010 filter
- 2009 (56) Apply 2009 filter
- 2008 (40) Apply 2008 filter
- 2007 (21) Apply 2007 filter
- 2006 (3) Apply 2006 filter
2454 Janelia Publications
Showing 2021-2030 of 2454 resultsTularemia is a deadly, febrile disease caused by infection by the gram-negative bacterium, Francisella tularensis. Members of the ubiquitous serine hydrolase protein family are among current targets to treat diverse bacterial infections. Herein we present a structural and functional study of a novel bacterial carboxylesterase (FTT258) from F. tularensis, a homologue of human acyl protein thioesterase (hAPT1). The structure of FTT258 has been determined in multiple forms, and unexpectedly large conformational changes of a peripheral flexible loop occur in the presence of a mechanistic cyclobutanone ligand. The concomitant changes in this hydrophobic loop and the newly exposed hydrophobic substrate binding pocket suggest that the observed structural changes are essential to the biological function and catalytic activity of FTT258. Using diverse substrate libraries, site-directed mutagenesis, and liposome binding assays, we determined the importance of these structural changes to the catalytic activity and membrane binding activity of FTT258. Residues within the newly exposed hydrophobic binding pocket and within the peripheral flexible loop proved essential to the hydrolytic activity of FTT258, indicating that structural rearrangement is required for catalytic activity. Both FTT258 and hAPT1 also showed significant association with liposomes designed to mimic bacterial or human membranes, respectively, even though similar structural rearrangements for hAPT1 have not been reported. The necessity for acyl protein thioesterases to have maximal catalytic activity near the membrane surface suggests that these conformational changes in the protein may dually regulate catalytic activity and membrane association in bacterial and human homologues.
Two clichés of science journalism have now played out around the ENCODE project. ENCODE’s publicity first presented a misleading "all the textbooks are wrong" narrative about noncoding human DNA. Now several critiques of ENCODE’s narrative have been published, and one was so vitriolic that it fueled "undignified academic squabble" stories that focused on tone more than substance. Neither story line does justice to our actual understanding of genomes, to ENCODE’s results, or to the role of big science in biology.
Electron cryomicroscopy, or cryoEM, is an emerging technique for studying the three-dimensional structures of proteins and large macromolecular machines. Electron crystallography is a branch of cryoEM in which structures of proteins can be studied at resolutions that rival those achieved by X-ray crystallography. Electron crystallography employs two-dimensional crystals of a membrane protein embedded within a lipid bilayer. The key to a successful electron crystallographic experiment is the crystallization, or reconstitution, of the protein of interest. This unit describes ways in which protein can be expressed, purified, and reconstituted into well-ordered two-dimensional crystals. A protocol is also provided for negative stain electron microscopy as a tool for screening crystallization trials. When large and well-ordered crystals are obtained, the structures of both protein and its surrounding membrane can be determined to atomic resolution.

Object detection and classification are key tasks in computer vision that can facilitate high-throughput image analysis of microscopy data. We present a set of local image descriptors for three-dimensional (3D) microscopy datasets inspired by the well-known Haar wavelet framework. We add orientation, illumination and scale information by assuming that the neighborhood surrounding points of interests in the image can be described with ellipsoids, and we increase discriminative power by incorporating edge and shape information into the features. The calculation of the local image descriptors is implemented in a Graphics Processing Unit (GPU) in order to reduce computation time to 1 millisecond per object of interest. We present results for cell division detection in 3D time-lapse fluorescence microscopy with 97.6% accuracy.

Understanding the development of complex multicellular organisms as a function of the underlying cell behavior is one of the most fundamental goals of developmental biology. The ability to quantitatively follow cell dynamics in entire developing embryos is an indispensable step towards such a system-level understanding. In recent years, light-sheet fluorescence microscopy has emerged as a particularly promising strategy for recording the in vivo data required to realize this goal. Using light-sheet fluorescence microscopy, entire complex organisms can be rapidly imaged in three dimensions at sub-cellular resolution, achieving high temporal sampling and excellent signal-to-noise ratio without damaging the living specimen or bleaching fluorescent markers. The resulting datasets allow following individual cells in vertebrate and higher invertebrate embryos over up to several days of development. However, the complexity and size of these multi-terabyte recordings typically preclude comprehensive manual analyses. Thus, new computational approaches are required to automatically segment cell morphologies, accurately track cell identities and systematically analyze cell behavior throughout embryonic development. We review current efforts in light-sheet microscopy and bioimage informatics towards this goal, and argue that comprehensive cell lineage reconstructions are finally within reach for many key model organisms, including fruit fly, zebrafish and mouse.
Objective: While the contribution of α-Synuclein to neurodegeneration in Parkinson’s disease is well accepted, the putative impact of its close homologue, β-Synuclein, is enigmatic. β-Synuclein is widely expressed throughout the central nervous system as is α-Synuclein, but the physiological functions of both proteins remain unknown. Recent findings supported the view that β-Synuclein can act as an ameliorating regulator of α-Synuclein-induced neurotoxicity, having neuroprotective rather than neurodegenerative capabilities, and being non-aggregating due to absence of most part of the aggregation-promoting NAC domain. However, a mutation of β-Synuclein linked to dementia with Lewy bodies rendered the protein neurotoxic in transgenic mice and fibrillation of β-Synuclein has been demonstrated in vitro. Methods / Results: Supporting the hypothesis that β-Synuclein can act as a neurodegeneration-inducing factor we now demonstrate that wild-type β-Synuclein is neurotoxic for cultured primary neurons. Furthermore, β-Synuclein formed proteinase K resistant aggregates in dopaminergic neurons in vivo, leading to pronounced and progressive neurodegeneration in rats. Expression of β-Synuclein caused mitochondrial fragmentation, but this fragmentation did not render mitochondria non-functional in terms of ion handling and respiration even in late stages of neurodegeneration. A comparison of the neurodegenerative effects induced by α-, β-, and γ-Synuclein revealed that β-Synuclein was eventually as neurotoxic as α-Synuclein for nigral dopaminergic neurons, while γ-Synuclein proved to be non-toxic and had very low aggregation propensity. Interpretation: Our results suggest that the role of β-Synuclein as a putative modulator of neuropathology in aggregopathies like Parkinson’s disease and dementia with Lewy bodies needs to be revisited. ANN NEUROL 2013. © 2013 American Neurological Association.
Neuroscience is at a crossroads. Great effort is being invested into deciphering specific neural interactions and circuits. At the same time, there exist few general theories or principles that explain brain function. We attribute this disparity, in part, to limitations in current methodologies. Traditional neurophysiological approaches record the activities of one neuron or a few neurons at a time. Neurochemical approaches focus on single neurotransmitters. Yet, there is an increasing realization that neural circuits operate at emergent levels, where the interactions between hundreds or thousands of neurons, utilizing multiple chemical transmitters, generate functional states. Brains function at the nanoscale, so tools to study brains must ultimately operate at this scale, as well. Nanoscience and nanotechnology are poised to provide a rich toolkit of novel methods to explore brain function by enabling simultaneous measurement and manipulation of activity of thousands or even millions of neurons. We and others refer to this goal as the Brain Activity Mapping Project. In this Nano Focus, we discuss how recent developments in nanoscale analysis tools and in the design and synthesis of nanomaterials have generated optical, electrical, and chemical methods that can readily be adapted for use in neuroscience. These approaches represent exciting areas of technical development and research. Moreover, unique opportunities exist for nanoscientists, nanotechnologists, and other physical scientists and engineers to contribute to tackling the challenging problems involved in understanding the fundamentals of brain function.
Developmental signals such as Wnts are often presented to cells in an oriented manner. To examine the consequences of local Wnt signaling, we immobilized Wnt proteins on beads and introduced them to embryonic stem cells in culture. At the single-cell level, the Wnt-bead induced asymmetric distribution of Wnt-β-catenin signaling components, oriented the plane of mitotic division, and directed asymmetric inheritance of centrosomes. Before cytokinesis was completed, the Wnt-proximal daughter cell expressed high levels of nuclear β-catenin and pluripotency genes, whereas the distal daughter cell acquired hallmarks of differentiation. We suggest that a spatially restricted Wnt signal induces an oriented cell division that generates distinct cell fates at predictable positions relative to the Wnt source.
Automatic 3D digital reconstruction (tracing) of neurons embedded in noisy microscopic images is challenging, especially when the cell morphology is complex.
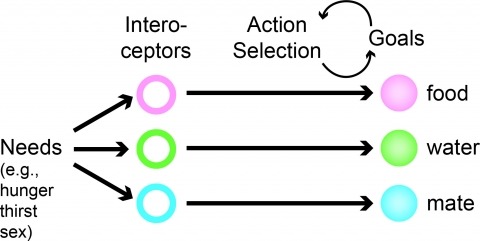
Neural processes that direct an animal’s actions toward environmental goals are critical elements for understanding behavior. The hypothalamus is closely associated with motivated behaviors required for survival and reproduction. Intense feeding, drinking, aggressive, and sexual behaviors can be produced by a simple neuronal stimulus applied to discrete hypothalamic regions. What can these "evoked behaviors" teach us about the neural processes that determine behavioral intent and intensity? Small populations of neurons sufficient to evoke a complex motivated behavior may be used as entry points to identify circuits that energize and direct behavior to specific goals. Here, I review recent applications of molecular genetic, optogenetic, and pharmacogenetic approaches that overcome previous limitations for analyzing anatomically complex hypothalamic circuits and their interactions with the rest of the brain. These new tools have the potential to bridge the gaps between neurobiological and psychological thinking about the mechanisms of complex motivated behavior.