Filter
Associated Lab
- Betzig Lab (2) Apply Betzig Lab filter
- Bock Lab (1) Apply Bock Lab filter
- Cardona Lab (1) Apply Cardona Lab filter
- Dickson Lab (3) Apply Dickson Lab filter
- Druckmann Lab (1) Apply Druckmann Lab filter
- Dudman Lab (3) Apply Dudman Lab filter
- Eddy/Rivas Lab (2) Apply Eddy/Rivas Lab filter
- Egnor Lab (1) Apply Egnor Lab filter
- Fetter Lab (2) Apply Fetter Lab filter
- Fitzgerald Lab (2) Apply Fitzgerald Lab filter
- Gonen Lab (2) Apply Gonen Lab filter
- Heberlein Lab (3) Apply Heberlein Lab filter
- Hess Lab (1) Apply Hess Lab filter
- Jayaraman Lab (1) Apply Jayaraman Lab filter
- Kainmueller Lab (1) Apply Kainmueller Lab filter
- Karpova Lab (1) Apply Karpova Lab filter
- Keleman Lab (2) Apply Keleman Lab filter
- Keller Lab (1) Apply Keller Lab filter
- Koay Lab (1) Apply Koay Lab filter
- Lavis Lab (3) Apply Lavis Lab filter
- Lippincott-Schwartz Lab (1) Apply Lippincott-Schwartz Lab filter
- Magee Lab (1) Apply Magee Lab filter
- Pavlopoulos Lab (2) Apply Pavlopoulos Lab filter
- Reiser Lab (2) Apply Reiser Lab filter
- Riddiford Lab (2) Apply Riddiford Lab filter
- Romani Lab (1) Apply Romani Lab filter
- Rubin Lab (2) Apply Rubin Lab filter
- Schreiter Lab (3) Apply Schreiter Lab filter
- Sgro Lab (1) Apply Sgro Lab filter
- Shroff Lab (1) Apply Shroff Lab filter
- Simpson Lab (1) Apply Simpson Lab filter
- Spruston Lab (4) Apply Spruston Lab filter
- Stern Lab (9) Apply Stern Lab filter
- Svoboda Lab (3) Apply Svoboda Lab filter
- Tervo Lab (1) Apply Tervo Lab filter
- Tjian Lab (5) Apply Tjian Lab filter
- Truman Lab (1) Apply Truman Lab filter
- Turaga Lab (1) Apply Turaga Lab filter
Publication Date
- December 2007 (12) Apply December 2007 filter
- November 2007 (9) Apply November 2007 filter
- October 2007 (12) Apply October 2007 filter
- September 2007 (8) Apply September 2007 filter
- August 2007 (7) Apply August 2007 filter
- July 2007 (16) Apply July 2007 filter
- June 2007 (5) Apply June 2007 filter
- May 2007 (7) Apply May 2007 filter
- April 2007 (8) Apply April 2007 filter
- March 2007 (7) Apply March 2007 filter
- February 2007 (3) Apply February 2007 filter
- January 2007 (12) Apply January 2007 filter
- Remove 2007 filter 2007
Type of Publication
106 Publications
Showing 41-50 of 106 resultsSpinocerebellar ataxia type-3 (SCA3) is among the most common dominantly inherited ataxias, and is one of nine devastating human neurodegenerative diseases caused by the expansion of a CAG repeat encoding glutamine within the gene. The polyglutamine domain confers toxicity on the protein Ataxin-3 leading to neuronal dysfunction and loss. Although modifiers of polyglutamine toxicity have been identified, little is known concerning how the modifiers function mechanistically to affect toxicity. To reveal insight into spinocerebellar ataxia type-3, we performed a genetic screen in Drosophila with pathogenic Ataxin-3-induced neurodegeneration and identified 25 modifiers defining 18 genes. Despite a variety of predicted molecular activities, biological analysis indicated that the modifiers affected protein misfolding. Detailed mechanistic studies revealed that some modifiers affected protein accumulation in a manner dependent on the proteasome, whereas others affected autophagy. Select modifiers of Ataxin-3 also affected tau, revealing common pathways between degeneration due to distinct human neurotoxic proteins. These findings provide new insight into molecular pathways of polyQ toxicity, defining novel targets for promoting neuronal survival in human neurodegenerative disease.
BACKGROUND: In many animals, the first few hours of life proceed with little or no transcription, and developmental regulation at these early stages is dependent on maternal cytoplasm rather than the zygotic nucleus. Translational control is critical for early Drosophila embryogenesis and is exerted mainly at the gene level. To understand post-transcriptional regulation during Drosophila early embryonic development, we used sucrose polysomal gradient analyses and GeneChip analysis to illustrate the translation profile of individual mRNAs. RESULTS: We determined ribosomal density and ribosomal occupancy of over 10,000 transcripts during the first ten hours after egg laying. CONCLUSION: We report the extent and general nature of gene regulation at the translational level during early Drosophila embryogenesis on a genome-wide basis. The diversity of the translation profiles indicates multiple mechanisms modulating transcript-specific translation. Cluster analyses suggest that the genes involved in some biological processes are co-regulated at the translational level at certain developmental stages.
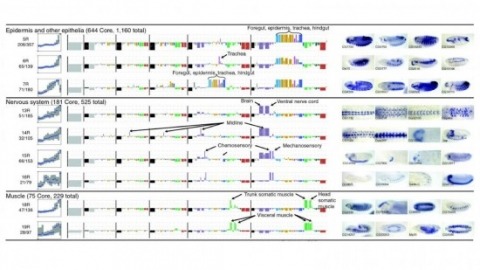
Cell and tissue specific gene expression is a defining feature of embryonic development in multi-cellular organisms. However, the range of gene expression patterns, the extent of the correlation of expression with function, and the classes of genes whose spatial expression are tightly regulated have been unclear due to the lack of an unbiased, genome-wide survey of gene expression patterns.
Habituation is a universal form of nonassociative learning that results in the devaluation of sensory inputs that have little information content. Although habituation is found throughout nature and has been studied in many organisms, the underlying molecular mechanisms remain poorly understood. We performed a forward genetic screen in Drosophila to search for mutations that modified habituation of an olfactory-mediated locomotor startle response, and we isolated a mutation in the glycogen synthase kinase-3 (GSK-3) homolog Shaggy. Decreases in Shaggy levels blunted habituation, whereas increases promoted habituation. Additionally, habituation acutely regulated Shaggy by an inhibitory phosphorylation mechanism, suggesting that a signal transduction pathway that regulates Shaggy is engaged during habituation. Although shaggy mutations also affected circadian rhythm period, this requirement was genetically separable from its role in habituation. Thus, shaggy functions in different neuronal circuits to regulate behavioral plasticity to an olfactory startle and circadian rhythmicity.
RATIONALE: This review provides insight for the judicious selection of nicotine dose ranges and routes of administration for in vivo studies. The literature is replete with reports in which a dosaging regimen chosen for a specific nicotine-mediated response was suboptimal for the species used. In many cases, such discrepancies could be attributed to the complex variables comprising species-specific in vivo responses to acute or chronic nicotine exposure. OBJECTIVES: This review capitalizes on the authors' collective decades of in vivo nicotine experimentation to clarify the issues and to identify the variables to be considered in choosing a dosaging regimen. Nicotine dose ranges tolerated by humans and their animal models provide guidelines for experiments intended to extrapolate to human tobacco exposure through cigarette smoking or nicotine replacement therapies. Just as important are the nicotine dosaging regimens used to provide a mechanistic framework for acquisition of drug-taking behavior, dependence, tolerance, or withdrawal in animal models. RESULTS: Seven species are addressed: humans, nonhuman primates, rats, mice, Drosophila, Caenorhabditis elegans, and zebrafish. After an overview on nicotine metabolism, each section focuses on an individual species, addressing issues related to genetic background, age, acute vs chronic exposure, route of administration, and behavioral responses. CONCLUSIONS: The selected examples of successful dosaging ranges are provided, while emphasizing the necessity of empirically determined dose-response relationships based on the precise parameters and conditions inherent to a specific hypothesis. This review provides a new, experimentally based compilation of species-specific dose selection for studies on the in vivo effects of nicotine.
HCN1 hyperpolarization-activated cation channels act as an inhibitory constraint of both spatial learning and synaptic integration and long-term plasticity in the distal dendrites of hippocampal CA1 pyramidal neurons. However, as HCN1 channels provide an excitatory current, the mechanism of their inhibitory action remains unclear. Here we report that HCN1 channels also constrain CA1 distal dendritic Ca2+ spikes, which have been implicated in the induction of LTP at distal excitatory synapses. Our experimental and computational results indicate that HCN1 channels provide both an active shunt conductance that decreases the temporal integration of distal EPSPs and a tonic depolarizing current that increases resting inactivation of T-type and N-type voltage-gated Ca2+ channels, which contribute to the Ca2+ spikes. This dual mechanism may provide a general means by which HCN channels regulate dendritic excitability.
Whereas recent studies have elucidated principles for representation of information within the entorhinal cortex, less is known about the molecular basis for information processing by entorhinal neurons. The HCN1 gene encodes ion channels that mediate hyperpolarization-activated currents (I(h)) that control synaptic integration and influence several forms of learning and memory. We asked whether hyperpolarization-activated, cation nonselective 1 (HCN1) channels control processing of information by stellate cells found within layer II of the entorhinal cortex. Axonal projections from these neurons form a major component of the synaptic input to the dentate gyrus of the hippocampus. To determine whether HCN1 channels control either the resting or the active properties of stellate neurons, we performed whole-cell recordings in horizontal brain slices prepared from adult wild-type and HCN1 knock-out mice. We found that HCN1 channels are required for rapid and full activation of hyperpolarization-activated currents in stellate neurons. HCN1 channels dominate the membrane conductance at rest, are not required for theta frequency (4-12 Hz) membrane potential fluctuations, but suppress low-frequency (<4 Hz) components of spontaneous and evoked membrane potential activity. During sustained activation of stellate cells sufficient for firing of repeated action potentials, HCN1 channels control the pattern of spike output by promoting recovery of the spike afterhyperpolarization. These data suggest that HCN1 channels expressed by stellate neurons in layer II of the entorhinal cortex are key molecular components in the processing of inputs to the hippocampal dentate gyrus, with distinct integrative roles during resting and active states.
Genetic studies of the targets of the Hox genes have revealed only the tip of the iceberg. Recent microarray studies that have identified hundreds more transcriptional responses to Hox genes in Drosophila will help elucidate the role of Hox genes in development and evolution.
Posttranslational modification through palmitoylation regulates protein localization and function. In this study, we identify a role for the Drosophila melanogaster palmitoyl transferase Huntingtin-interacting protein 14 (HIP14) in neurotransmitter release. hip14 mutants show exocytic defects at low frequency stimulation and a nearly complete loss of synaptic transmission at higher temperature. Interestingly, two exocytic components known to be palmitoylated, cysteine string protein (CSP) and SNAP25, are severely mislocalized at hip14 mutant synapses. Complementary DNA rescue and localization experiments indicate that HIP14 is required solely in the nervous system and is essential for presynaptic function. Biochemical studies indicate that HIP14 palmitoylates CSP and that CSP is not palmitoylated in hip14 mutants. Furthermore, the hip14 exocytic defects can be suppressed by targeting CSP to synaptic vesicles using a chimeric protein approach. Our data indicate that HIP14 controls neurotransmitter release by regulating the trafficking of CSP to synapses.
We present hysteresis phenomena of the intelligent driver model for traffic flow in a circular one-lane roadway. We show that the microscopic structure of traffic flow is dependent on its initial state by plotting the fraction of congested vehicles over the density, which shows a typical hysteresis loop, and by investigating the trajectories of vehicles on the velocity-over-headway plane. We find that the trajectories of vehicles on the velocity-over-headway plane, which usually show a hysteresis loop, include multiple loops. We also point out the relations between these hysteresis loops and the congested jams or high-density clusters in traffic flow.