Filter
Associated Lab
- Ahrens Lab (5) Apply Ahrens Lab filter
- Aso Lab (1) Apply Aso Lab filter
- Baker Lab (2) Apply Baker Lab filter
- Betzig Lab (4) Apply Betzig Lab filter
- Bock Lab (2) Apply Bock Lab filter
- Cardona Lab (1) Apply Cardona Lab filter
- Cui Lab (2) Apply Cui Lab filter
- Dickson Lab (3) Apply Dickson Lab filter
- Druckmann Lab (1) Apply Druckmann Lab filter
- Dudman Lab (2) Apply Dudman Lab filter
- Eddy/Rivas Lab (2) Apply Eddy/Rivas Lab filter
- Egnor Lab (1) Apply Egnor Lab filter
- Fetter Lab (3) Apply Fetter Lab filter
- Fitzgerald Lab (1) Apply Fitzgerald Lab filter
- Gonen Lab (11) Apply Gonen Lab filter
- Grigorieff Lab (4) Apply Grigorieff Lab filter
- Harris Lab (3) Apply Harris Lab filter
- Heberlein Lab (6) Apply Heberlein Lab filter
- Hermundstad Lab (1) Apply Hermundstad Lab filter
- Hess Lab (2) Apply Hess Lab filter
- Jayaraman Lab (3) Apply Jayaraman Lab filter
- Ji Lab (1) Apply Ji Lab filter
- Johnson Lab (1) Apply Johnson Lab filter
- Karpova Lab (1) Apply Karpova Lab filter
- Keller Lab (10) Apply Keller Lab filter
- Lavis Lab (4) Apply Lavis Lab filter
- Leonardo Lab (3) Apply Leonardo Lab filter
- Lippincott-Schwartz Lab (11) Apply Lippincott-Schwartz Lab filter
- Looger Lab (10) Apply Looger Lab filter
- Magee Lab (3) Apply Magee Lab filter
- Menon Lab (3) Apply Menon Lab filter
- Pachitariu Lab (3) Apply Pachitariu Lab filter
- Pavlopoulos Lab (1) Apply Pavlopoulos Lab filter
- Reiser Lab (2) Apply Reiser Lab filter
- Riddiford Lab (5) Apply Riddiford Lab filter
- Romani Lab (1) Apply Romani Lab filter
- Rubin Lab (5) Apply Rubin Lab filter
- Satou Lab (2) Apply Satou Lab filter
- Scheffer Lab (3) Apply Scheffer Lab filter
- Schreiter Lab (7) Apply Schreiter Lab filter
- Sgro Lab (1) Apply Sgro Lab filter
- Singer Lab (9) Apply Singer Lab filter
- Spruston Lab (2) Apply Spruston Lab filter
- Stern Lab (6) Apply Stern Lab filter
- Sternson Lab (3) Apply Sternson Lab filter
- Svoboda Lab (10) Apply Svoboda Lab filter
- Tjian Lab (1) Apply Tjian Lab filter
- Truman Lab (3) Apply Truman Lab filter
- Turaga Lab (2) Apply Turaga Lab filter
- Turner Lab (2) Apply Turner Lab filter
- Wu Lab (3) Apply Wu Lab filter
- Zlatic Lab (2) Apply Zlatic Lab filter
Associated Project Team
Publication Date
- December 2013 (13) Apply December 2013 filter
- November 2013 (10) Apply November 2013 filter
- October 2013 (20) Apply October 2013 filter
- September 2013 (19) Apply September 2013 filter
- August 2013 (15) Apply August 2013 filter
- July 2013 (19) Apply July 2013 filter
- June 2013 (17) Apply June 2013 filter
- May 2013 (10) Apply May 2013 filter
- April 2013 (12) Apply April 2013 filter
- March 2013 (11) Apply March 2013 filter
- February 2013 (19) Apply February 2013 filter
- January 2013 (29) Apply January 2013 filter
- Remove 2013 filter 2013
Type of Publication
194 Publications
Showing 61-70 of 194 resultsGap junctions (GJs) represent connexin-rich membrane domains that connect interiors of adjoining cells in mammalian tissues. How fast GJs can respond to bacterial pathogens has not been known previously. Using Bessel beam plane illumination and confocal spinning disk microscopy, we found fast (~500 ms) formation of connexin-depleted regions (CDRs) inside GJ plaques between cells exposed to AB5 toxins. CDR formation appears as a fast redistribution of connexin channels within GJ plaques with minor changes in outline or geometry. CDR formation does not depend on membrane trafficking or submembrane cytoskeleton and has no effect on GJ conductance. However, CDR responses depend on membrane lipids, can be modified by cholesterol-clustering agents and extracellular K(+) ion concentration, and influence cAMP signaling. The CDR response of GJ plaques to bacterial toxins is a phenomenon observed for all tested connexin isoforms. Through signaling, the CDR response may enable cells to sense exposure to AB5 toxins. CDR formation may reflect lipid-phase separation events in the biological membrane of the GJ plaque, leading to increased connexin packing and lipid reorganization. Our data demonstrate very fast dynamics (in the millisecond-to-second range) within GJ plaques, which previously were considered to be relatively stable, long-lived structures.
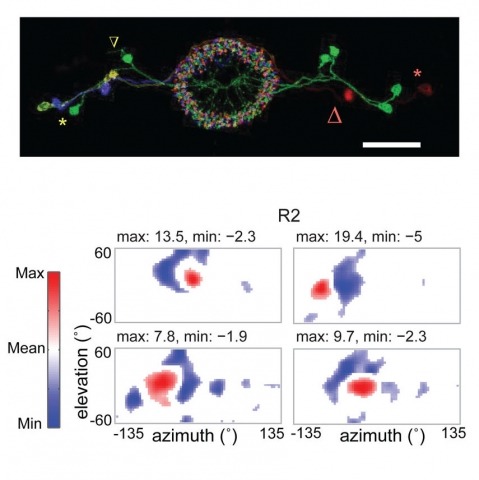
Many animals, including insects, are known to use visual landmarks to orient in their environment. In Drosophila melanogaster, behavioural genetics studies have identified a higher brain structure called the central complex as being required for the fly’s innate responses to vertical visual features and its short- and long-term memory for visual patterns. But whether and how neurons of the fly central complex represent visual features are unknown. Here we use two-photon calcium imaging in head-fixed walking and flying flies to probe visuomotor responses of ring neurons—a class of central complex neurons that have been implicated in landmark-driven spatial memory in walking flies and memory for visual patterns in tethered flying flies. We show that dendrites of ring neurons are visually responsive and arranged retinotopically. Ring neuron receptive fields comprise both excitatory and inhibitory subfields, resembling those of simple cells in the mammalian primary visual cortex. Ring neurons show strong and, in some cases, direction-selective orientation tuning, with a notable preference for vertically oriented features similar to those that evoke innate responses in flies. Visual responses were diminished during flight, but, in contrast with the hypothesized role of the central complex in the control of locomotion, not modulated during walking. Taken together, these results indicate that ring neurons represent behaviourally relevant visual features in the fly’s environment, enabling downstream central complex circuits to produce appropriate motor commands. More broadly, this study opens the door to mechanistic investigations of circuit computations underlying visually guided action selection in the Drosophila central complex.
In vertebrates, primary sex determination refers to the decision within a bipotential organ precursor to differentiate as a testis or ovary. Bifurcation of organ fate begins between embryonic day (E) 11.0–E12.0 in mice and likely involves a dynamic transcription network that is poorly understood. To elucidate the first steps of sexual fate specification, we profiled the XX and XY gonad transcriptomes at fine granularity during this period and resolved cascades of gene activation and repression. C57BL/6J (B6) XY gonads showed a consistent 5-hour delay in the activation of most male pathway genes and repression of female pathway genes relative to 129S1/SvImJ, which likely explains the sensitivity of the B6 strain to male-to-female sex reversal. Using this fine time course data, we predicted novel regulatory genes underlying expression QTLs (eQTLs) mapped in a previous study. To test predictions, we developed an in vitro gonad primary cell assay and optimized a lentivirus-based shRNA delivery method to silence candidate genes and quantify effects on putative targets. We provide strong evidence that Lmo4 (Lim-domain only 4) is a novel regulator of sex determination upstream of SF1 (Nr5a1), Sox9, Fgf9, and Col9a3. This approach can be readily applied to identify regulatory interactions in other systems.
Identifying the neuronal ensembles that respond to specific stimuli and mapping their projection patterns in living animals are fundamental challenges in neuroscience. To this end, we engineered a synthetic promoter, the enhanced synaptic activity-responsive element (E-SARE), that drives neuronal activity-dependent gene expression more potently than other existing immediate-early gene promoters. Expression of a drug-inducible Cre recombinase downstream of E-SARE enabled imaging of neuronal populations that respond to monocular visual stimulation and tracking of their long-distance thalamocortical projections in living mice. Targeted cell-attached recordings and calcium imaging of neurons in sensory cortices revealed that E-SARE reporter expression correlates with sensory-evoked neuronal activity at the single-cell level and is highly specific to the type of stimuli presented to the animals. This activity-dependent promoter can expand the repertoire of genetic approaches for high-resolution anatomical and functional analysis of neural circuits.
Genetically hard-wired neural mechanisms must enforce behavioral reproductive isolation because interspecies courtship is rare even in sexually na{\"ıve animals of most species. We find that the chemoreceptor Gr32a inhibits male D. melanogaster from courting diverse fruit fly species. Gr32a recognizes nonvolatile aversive cues present on these reproductively dead-end targets, and activity of Gr32a neurons is necessary and sufficient to inhibit interspecies courtship. Male-specific Fruitless (Fru(M)), a master regulator of courtship, also inhibits interspecies courtship. Gr32a and Fru(M) are not coexpressed, but Fru(M) neurons contact Gr32a neurons, suggesting that these genes influence a shared neural circuit that inhibits interspecies courtship. Gr32a and Fru(M) also suppress within-species intermale courtship, but we show that distinct mechanisms preclude sexual displays toward conspecific males and other species. Although this chemosensory pathway does not inhibit interspecies mating in D. melanogaster females, similar mechanisms appear to inhibit this behavior in many other male drosophilids.
The discovery of intracellular Ca(2+) signals within astrocytes has changed our view of how these ubiquitous cells contribute to brain function. Classically thought merely to serve supportive functions, astrocytes are increasingly thought to respond to, and regulate, neurons. The use of organic Ca(2+) indicator dyes such as Fluo-4 and Fura-2 has proved instrumental in the study of astrocyte physiology. However, progress has recently been accelerated by the use of cytosolic and membrane targeted genetically encoded calcium indicators (GECIs). Herein, we review these recent findings, discuss why studying astrocyte Ca(2+) signals is important to understand brain function, and summarize work that led to the discovery of TRPA1 channel-mediated near-membrane Ca(2+) signals in astrocytes and their indirect neuromodulatory roles at inhibitory synapses in the CA1 stratum radiatum region of the hippocampus. We suggest that the use of membrane-targeted and cytosolic GECIs holds great promise to explore the diversity of Ca(2+) signals within single astrocytes and also to study diversity of function for astrocytes in different parts of the brain.
Genetically encoded calcium indicators (GECIs) are powerful tools for systems neuroscience. Here we describe red, single-wavelength GECIs, "RCaMPs," engineered from circular permutation of the thermostable red fluorescent protein mRuby. High-resolution crystal structures of mRuby, the red sensor RCaMP, and the recently published red GECI R-GECO1 give insight into the chromophore environments of the Ca(2+)-bound state of the sensors and the engineered protein domain interfaces of the different indicators. We characterized the biophysical properties and performance of RCaMP sensors in vitro and in vivo in Caenorhabditis elegans, Drosophila larvae, and larval zebrafish. Further, we demonstrate 2-color calcium imaging both within the same cell (registering mitochondrial and somatic [Ca(2+)]) and between two populations of cells: neurons and astrocytes. Finally, we perform integrated optogenetics experiments, wherein neural activation via channelrhodopsin-2 (ChR2) or a red-shifted variant, and activity imaging via RCaMP or GCaMP, are conducted simultaneously, with the ChR2/RCaMP pair providing independently addressable spectral channels. Using this paradigm, we measure calcium responses of naturalistic and ChR2-evoked muscle contractions in vivo in crawling C. elegans. We systematically compare the RCaMP sensors to R-GECO1, in terms of action potential-evoked fluorescence increases in neurons, photobleaching, and photoswitching. R-GECO1 displays higher Ca(2+) affinity and larger dynamic range than RCaMP, but exhibits significant photoactivation with blue and green light, suggesting that integrated channelrhodopsin-based optogenetics using R-GECO1 may be subject to artifact. Finally, we create and test blue, cyan, and yellow variants engineered from GCaMP by rational design. This engineered set of chromatic variants facilitates new experiments in functional imaging and optogenetics.
Secretion systems require high-fidelity mechanisms to discriminate substrates among the vast cytoplasmic pool of proteins. Factors mediating substrate recognition by the type VI secretion system (T6SS) of Gram-negative bacteria, a widespread pathway that translocates effector proteins into target bacterial cells, have not been defined. We report that haemolysin coregulated protein (Hcp), a ring-shaped hexamer secreted by all characterized T6SSs, binds specifically to cognate effector molecules. Electron microscopy analysis of an Hcp-effector complex from Pseudomonas aeruginosa revealed the effector bound to the inner surface of Hcp. Further studies demonstrated that interaction with the Hcp pore is a general requirement for secretion of diverse effectors encompassing several enzymatic classes. Though previous models depict Hcp as a static conduit, our data indicate it is a chaperone and receptor of substrates. These unique functions of a secreted protein highlight fundamental differences between the export mechanism of T6 and other characterized secretory pathways.
The internal ribosome entry site (IRES) of the hepatitis C virus (HCV) drives noncanonical initiation of protein synthesis necessary for viral replication. Functional studies of the HCV IRES have focused on 80S ribosome formation but have not explored its role after the 80S ribosome is poised at the start codon. Here, we report that mutations of an IRES domain that docks in the 40S subunit’s decoding groove cause only a local perturbation in IRES structure and result in conformational changes in the IRES-rabbit 40S subunit complex. Functionally, the mutations decrease IRES activity by inhibiting the first ribosomal translocation event, and modeling results suggest that this effect occurs through an interaction with a single ribosomal protein. The ability of the HCV IRES to manipulate the ribosome provides insight into how the ribosome’s structure and function can be altered by bound RNAs, including those derived from cellular invaders.
Random scattering and aberrations severely limit the imaging depth in optical microscopy. We introduce a rapid, parallel wavefront compensation technique that efficiently compensates even highly complex phase distortions. Using coherence gated backscattered light as a feedback signal, we focus light deep inside highly scattering brain tissue. We demonstrate that the same wavefront optimization technique can also be used to compensate spectral phase distortions in ultrashort laser pulses using nonlinear iterative feedback. We can restore transform limited pulse durations at any selected target location and compensate for dispersion that has occurred in the optical train and within the sample.