Filter
Associated Lab
Associated Project Team
Publication Date
- 2015 (2) Apply 2015 filter
- 2014 (1) Apply 2014 filter
- 2011 (3) Apply 2011 filter
- 2010 (1) Apply 2010 filter
- 2009 (2) Apply 2009 filter
- 2008 (1) Apply 2008 filter
- 2006 (3) Apply 2006 filter
- 2005 (1) Apply 2005 filter
- 2004 (1) Apply 2004 filter
- 2003 (2) Apply 2003 filter
- 2002 (1) Apply 2002 filter
- 2001 (3) Apply 2001 filter
- 2000 (3) Apply 2000 filter
- 1985 (1) Apply 1985 filter
Type of Publication
25 Publications
Showing 21-25 of 25 resultsThe sense of taste provides animals with valuable information about the nature and quality of food. Bitter taste detection functions as an important sensory input to warn against the ingestion of toxic and noxious substances. T2Rs are a family of approximately 30 highly divergent G-protein-coupled receptors (GPCRs) that are selectively expressed in the tongue and palate epithelium and are implicated in bitter taste sensing. Here we demonstrate, using a combination of genetic, behavioural and physiological studies, that T2R receptors are necessary and sufficient for the detection and perception of bitter compounds, and show that differences in T2Rs between species (human and mouse) can determine the selectivity of bitter taste responses. In addition, we show that mice engineered to express a bitter taste receptor in ’sweet cells’ become strongly attracted to its cognate bitter tastants, whereas expression of the same receptor (or even a novel GPCR) in T2R-expressing cells resulted in mice that are averse to the respective compounds. Together these results illustrate the fundamental principle of bitter taste coding at the periphery: dedicated cells act as broadly tuned bitter sensors that are wired to mediate behavioural aversion.
Sweet and umami (the taste of monosodium glutamate) are the main attractive taste modalities in humans. T1Rs are candidate mammalian taste receptors that combine to assemble two heteromeric G-protein-coupled receptor complexes: T1R1+3, an umami sensor, and T1R2+3, a sweet receptor. We now report the behavioral and physiological characterization of T1R1, T1R2, and T1R3 knockout mice. We demonstrate that sweet and umami taste are strictly dependent on T1R-receptors, and show that selective elimination of T1R-subunits differentially abolishes detection and perception of these two taste modalities. To examine the basis of sweet tastant recognition and coding, we engineered animals expressing either the human T1R2-receptor (hT1R2), or a modified opioid-receptor (RASSL) in sweet cells. Expression of hT1R2 in mice generates animals with humanized sweet taste preferences, while expression of RASSL drives strong attraction to a synthetic opiate, demonstrating that sweet cells trigger dedicated behavioral outputs, but their tastant selectivity is determined by the nature of the receptors.
Carbonated beverages are commonly available and immensely popular, but little is known about the cellular and molecular mechanisms underlying the perception of carbonation in the mouth. In mammals, carbonation elicits both somatosensory and chemosensory responses, including activation of taste neurons. We have identified the cellular and molecular substrates for the taste of carbonation. By targeted genetic ablation and the silencing of synapses in defined populations of taste receptor cells, we demonstrated that the sour-sensing cells act as the taste sensors for carbonation, and showed that carbonic anhydrase 4, a glycosylphosphatidylinositol-anchored enzyme, functions as the principal CO2 taste sensor. Together, these studies reveal the basis of the taste of carbonation as well as the contribution of taste cells in the orosensory response to CO2.
Eyes differ markedly in the animal kingdom, and are an extreme example of the evolution of multiple anatomical solutions to light detection and image formation. A salient feature of all photoreceptor cells is the presence of a specialized compartment (disc outer segments in vertebrates, and microvillar rhabdomeres in insects), whose primary role is to accommodate the millions of light receptor molecules required for efficient photon collection. In insects, compound eyes can have very different inner architectures. Fruitflies and houseflies have an open rhabdom system, in which the seven rhabdomeres of each ommatidium are separated from each other and function as independent light guides. In contrast, bees and various mosquitoes and beetle species have a closed system, in which rhabdomeres within each ommatidium are fused to each other, thus sharing the same visual axis. To understand the transition between open and closed rhabdom systems, we isolated and characterized the role of Drosophila genes involved in rhabdomere assembly. Here we show that Spacemaker, a secreted protein expressed only in the eyes of insects with open rhabdom systems, acts together with Prominin and the cell adhesion molecule Chaoptin to choreograph the partitioning of rhabdomeres into an open system. Furthermore, the complete loss of spacemaker (spam) converts an open rhabdom system to a closed one, whereas its targeted expression to photoreceptors of a closed system markedly reorganizes the architecture of the compound eyes to resemble an open system. Our results provide a molecular atlas for the construction of microvillar assemblies and illustrate the critical effect of differences in a single structural protein in morphogenesis.
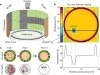
The ability of insects to learn and navigate to specific locations in the environment has fascinated naturalists for decades. The impressive navigational abilities of ants, bees, wasps and other insects demonstrate that insects are capable of visual place learning, but little is known about the underlying neural circuits that mediate these behaviours. Drosophila melanogaster (common fruit fly) is a powerful model organism for dissecting the neural circuitry underlying complex behaviours, from sensory perception to learning and memory. Drosophila can identify and remember visual features such as size, colour and contour orientation. However, the extent to which they use vision to recall specific locations remains unclear. Here we describe a visual place learning platform and demonstrate that Drosophila are capable of forming and retaining visual place memories to guide selective navigation. By targeted genetic silencing of small subsets of cells in the Drosophila brain, we show that neurons in the ellipsoid body, but not in the mushroom bodies, are necessary for visual place learning. Together, these studies reveal distinct neuroanatomical substrates for spatial versus non-spatial learning, and establish Drosophila as a powerful model for the study of spatial memories.