Filter
Associated Lab
- Ahrens Lab (3) Apply Ahrens Lab filter
- Betzig Lab (8) Apply Betzig Lab filter
- Card Lab (2) Apply Card Lab filter
- Cardona Lab (1) Apply Cardona Lab filter
- Druckmann Lab (1) Apply Druckmann Lab filter
- Eddy/Rivas Lab (2) Apply Eddy/Rivas Lab filter
- Fetter Lab (4) Apply Fetter Lab filter
- Fitzgerald Lab (1) Apply Fitzgerald Lab filter
- Gonen Lab (6) Apply Gonen Lab filter
- Harris Lab (1) Apply Harris Lab filter
- Heberlein Lab (3) Apply Heberlein Lab filter
- Hess Lab (2) Apply Hess Lab filter
- Ji Lab (4) Apply Ji Lab filter
- Kainmueller Lab (1) Apply Kainmueller Lab filter
- Keller Lab (3) Apply Keller Lab filter
- Lavis Lab (4) Apply Lavis Lab filter
- Looger Lab (3) Apply Looger Lab filter
- Magee Lab (4) Apply Magee Lab filter
- Murphy Lab (1) Apply Murphy Lab filter
- Pastalkova Lab (3) Apply Pastalkova Lab filter
- Reiser Lab (1) Apply Reiser Lab filter
- Riddiford Lab (5) Apply Riddiford Lab filter
- Romani Lab (2) Apply Romani Lab filter
- Rubin Lab (4) Apply Rubin Lab filter
- Saalfeld Lab (2) Apply Saalfeld Lab filter
- Satou Lab (1) Apply Satou Lab filter
- Schreiter Lab (3) Apply Schreiter Lab filter
- Sgro Lab (1) Apply Sgro Lab filter
- Shroff Lab (6) Apply Shroff Lab filter
- Spruston Lab (3) Apply Spruston Lab filter
- Stern Lab (4) Apply Stern Lab filter
- Sternson Lab (1) Apply Sternson Lab filter
- Svoboda Lab (6) Apply Svoboda Lab filter
- Tjian Lab (7) Apply Tjian Lab filter
- Truman Lab (2) Apply Truman Lab filter
- Turner Lab (1) Apply Turner Lab filter
- Wu Lab (1) Apply Wu Lab filter
- Zuker Lab (1) Apply Zuker Lab filter
Publication Date
- December 2008 (11) Apply December 2008 filter
- November 2008 (8) Apply November 2008 filter
- October 2008 (7) Apply October 2008 filter
- September 2008 (12) Apply September 2008 filter
- August 2008 (10) Apply August 2008 filter
- July 2008 (14) Apply July 2008 filter
- June 2008 (13) Apply June 2008 filter
- May 2008 (8) Apply May 2008 filter
- April 2008 (7) Apply April 2008 filter
- March 2008 (16) Apply March 2008 filter
- February 2008 (13) Apply February 2008 filter
- January 2008 (21) Apply January 2008 filter
- Remove 2008 filter 2008
Type of Publication
140 Publications
Showing 131-140 of 140 results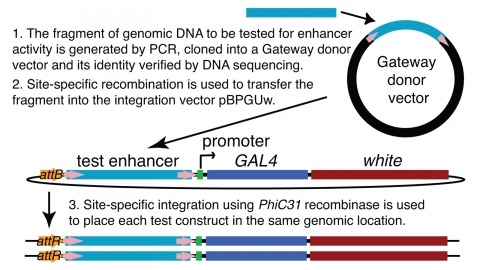
We demonstrate the feasibility of generating thousands of transgenic Drosophila melanogaster lines in which the expression of an exogenous gene is reproducibly directed to distinct small subsets of cells in the adult brain. We expect the expression patterns produced by the collection of 5,000 lines that we are currently generating to encompass all neurons in the brain in a variety of intersecting patterns. Overlapping 3-kb DNA fragments from the flanking noncoding and intronic regions of genes thought to have patterned expression in the adult brain were inserted into a defined genomic location by site-specific recombination. These fragments were then assayed for their ability to function as transcriptional enhancers in conjunction with a synthetic core promoter designed to work with a wide variety of enhancer types. An analysis of 44 fragments from four genes found that >80% drive expression patterns in the brain; the observed patterns were, on average, comprised of <100 cells. Our results suggest that the D. melanogaster genome contains >50,000 enhancers and that multiple enhancers drive distinct subsets of expression of a gene in each tissue and developmental stage. We expect that these lines will be valuable tools for neuroanatomy as well as for the elucidation of neuronal circuits and information flow in the fly brain.
p-Nitrophenyl acetate is the most commonly used substrate for detecting the catalytic activity of esterases, including those that activate prodrugs in human cells. This substrate is unstable in aqueous solution, limiting its utility. Here, a stable chromogenic substrate for esterases is produced by the structural isolation of an acetyl ester and p-nitroaniline group using a trimethyl lock moiety. Upon ester hydrolysis, unfavorable steric interactions between the three methyl groups of this o-hydroxycinnamic acid derivative encourage rapid lactonization to form a hydrocoumarin and release p-nitroaniline. This "prochromophore" could find use in a variety of assays.
In practice, understanding the spatial relationships between the surfaces of an object, can significantly improve the performance of object recognition systems. In this paper we propose a novel framework to recognize objects in pictures taken from arbitrary viewpoints. The idea is to maintain the frontal views of the major faces of objects in a global flat map. Then an unfolding warping technique is used to change the pose of the query object in the test view so that all visible surfaces of the object can be observed from a frontal viewpoint, improving the handling of serious occlusions and large viewpoint changes. We demonstrate the effectiveness of our approach through analysis of recognition trials of complex objects with comparison to popular methods.
Macaque monkeys were tested on a delayed-match-to-multiple-sample task, with either a limited set of well trained images (in randomized sequence) or with never-before-seen images. They performed much better with novel images. False positives were mostly limited to catch-trial image repetitions from the preceding trial. This result implies extremely effective one-shot learning, resembling Standing's finding that people detect familiarity for 10,000 once-seen pictures (with 80% accuracy) (Standing, 1973). Familiarity memory may differ essentially from identification, which embeds and generates contextual information. When encountering another person, we can say immediately whether his or her face is familiar. However, it may be difficult for us to identify the same person. To accompany the psychophysical findings, we present a generic neural network model reproducing these behaviors, based on the same conservative Hebbian synaptic plasticity that generates delay activity identification memory. Familiarity becomes the first step toward establishing identification. Adding an inter-trial reset mechanism limits false positives for previous-trial images. The model, unlike previous proposals, relates repetition-recognition with enhanced neural activity, as recently observed experimentally in 92% of differential cells in prefrontal cortex, an area directly involved in familiarity recognition. There may be an essential functional difference between enhanced responses to novel versus to familiar images: The maximal signal from temporal cortex is for novel stimuli, facilitating additional sensory processing of newly acquired stimuli. The maximal signal for familiar stimuli arising in prefrontal cortex facilitates the formation of selective delay activity, as well as additional consolidation of the memory of the image in an upstream cortical module.
The p2 progenitor domain in the ventral spinal cord gives rise to two interneuron subtypes: V2a and V2b. Delta-Notch-mediated cell-cell interactions between postmitotic immature neurons have been implicated in the segregation of neuron subtypes. However, lineage relationships between V2a and V2b neurons have not been reported. We address this issue using Tg[vsx1:GFP]zebrafish, a model system in which high GFP expression is initiated near the final stage of p2 progenitors. Cell fates were followed in progeny using time-lapse microscopy. Results indicate that the vast majority, if not all, of GFP-labeled p2 progenitors divide once to produce V2a/V2b neuron pairs,indicating that V2a and V2b neurons are generated by the asymmetric division of pair-producing progenitor cells. Together with evidence that Notch signaling is involved in the cell fate specification process, our results strongly suggest that Delta-Notch interactions between sister cells play a crucial role in the final outcome of these asymmetric divisions. This mechanism for determining cell fate is similar to asymmetric divisions that occur during Drosophila neurogenesis, where ganglion mother cells divide once to produce distinct neurons. However, unlike in Drosophila, the divisional axes of p2 progenitors in zebrafish were not fixed. We report that the terminal division of pair-producing progenitor cells in vertebrate neurogenesis can reproducibly produce two distinct neurons through a mechanism that may not depend on the orientation of the division axis.
The conditional expression of hairpin constructs in Drosophila melanogaster has emerged in recent years as a method of choice in functional genomic studies. To date, upstream activating site-driven RNA interference constructs have been inserted into the genome randomly using P-element-mediated transformation, which can result in false negatives due to variable expression. To avoid this problem, we have developed a transgenic RNA interference vector based on the phiC31 site-specific integration method.
The role of the foraging (for) gene, which encodes a cyclic guanosine-3’,5’-monophosphate (cGMP)-dependent protein kinase (PKG), in food-search behavior in Drosophila has been intensively studied. However, its functions in other complex behaviors have not been well-characterized. Here, we show experimentally in Drosophila that the for gene is required in the operant visual learning paradigm. Visual pattern memory was normal in a natural variant rover (for(R)) but was impaired in another natural variant sitter (for(S)), which has a lower PKG level. Memory defects in for(S) flies could be rescued by either constitutive or adult-limited expression of for in the fan-shaped body. Interestingly, we showed that such rescue also occurred when for was expressed in the ellipsoid body. Additionally, expression of for in the fifth layer of the fan-shaped body restored sufficient memory for the pattern parameter "elevation" but not for "contour orientation," whereas expression of for in the ellipsoid body restored sufficient memory for both parameters. Our study defines a Drosophila model for further understanding the role of cGMP-PKG signaling in associative learning/memory and the neural circuit underlying this for-dependent visual pattern memory.
A key feature of reactive behaviors is the ability to spatially localize a salient stimulus and act accordingly. Such sensory-motor transformations must be particularly fast and well tuned in escape behaviors, in which both the speed and accuracy of the evasive response determine whether an animal successfully avoids predation [1]. We studied the escape behavior of the fruit fly, Drosophila, and found that flies can use visual information to plan a jump directly away from a looming threat. This is surprising, given the architecture of the pathway thought to mediate escape [2, 3]. Using high-speed videography, we found that approximately 200 ms before takeoff, flies begin a series of postural adjustments that determine the direction of their escape. These movements position their center of mass so that leg extension will push them away from the expanding visual stimulus. These preflight movements are not the result of a simple feed-forward motor program because their magnitude and direction depend on the flies’ initial postural state. Furthermore, flies plan a takeoff direction even in instances when they choose not to jump. This sophisticated motor program is evidence for a form of rapid, visually mediated motor planning in a genetically accessible model organism.

For sensory signals to control an animal’s behavior, they must first be transformed into a format appropriate for use by its motor systems. This fundamental problem is faced by all animals, including humans. Beyond simple reflexes, little is known about how such sensorimotor transformations take place. Here we describe how the outputs of a well-characterized population of fly visual interneurons, lobula plate tangential cells (LPTCs), are used by the animal’s gaze-stabilizing neck motor system. The LPTCs respond to visual input arising from both self-rotations and translations of the fly. The neck motor system however is involved in gaze stabilization and thus mainly controls compensatory head rotations. We investigated how the neck motor system is able to selectively extract rotation information from the mixed responses of the LPTCs. We recorded extracellularly from fly neck motor neurons (NMNs) and mapped the directional preferences across their extended visual receptive fields. Our results suggest that-like the tangential cells-NMNs are tuned to panoramic retinal image shifts, or optic flow fields, which occur when the fly rotates about particular body axes. In many cases, tangential cells and motor neurons appear to be tuned to similar axes of rotation, resulting in a correlation between the coordinate systems the two neural populations employ. However, in contrast to the primarily monocular receptive fields of the tangential cells, most NMNs are sensitive to visual motion presented to either eye. This results in the NMNs being more selective for rotation than the LPTCs. Thus, the neck motor system increases its rotation selectivity by a comparatively simple mechanism: the integration of binocular visual motion information.
Widefield fluorescence microscopy is seeing dramatic improvements in resolution, reaching today 100 nm in all three dimensions. This gain in resolution is achieved by dispensing with uniform Köhler illumination. Instead, non-uniform excitation light patterns with sinusoidal intensity variations in one, two, or three dimensions are applied combined with powerful image reconstruction techniques. Taking advantage of non-linear fluorophore response to the excitation field, the resolution can be further improved down to several 10 nm. In this review article, we describe the image formation in the microscope and computational reconstruction of the high-resolution dataset when exciting the specimen with a harmonic light pattern conveniently generated by interfering laser beams forming standing waves. We will also discuss extensions to total internal reflection microscopy, non-linear microscopy, and three-dimensional imaging.