Filter
Associated Lab
- Aguilera Castrejon Lab (2) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (5) Apply Ahrens Lab filter
- Aso Lab (3) Apply Aso Lab filter
- Betzig Lab (7) Apply Betzig Lab filter
- Beyene Lab (1) Apply Beyene Lab filter
- Bock Lab (5) Apply Bock Lab filter
- Branson Lab (3) Apply Branson Lab filter
- Card Lab (2) Apply Card Lab filter
- Cardona Lab (4) Apply Cardona Lab filter
- Clapham Lab (2) Apply Clapham Lab filter
- Darshan Lab (2) Apply Darshan Lab filter
- Dickson Lab (5) Apply Dickson Lab filter
- Druckmann Lab (3) Apply Druckmann Lab filter
- Dudman Lab (4) Apply Dudman Lab filter
- Espinosa Medina Lab (3) Apply Espinosa Medina Lab filter
- Feliciano Lab (1) Apply Feliciano Lab filter
- Fitzgerald Lab (2) Apply Fitzgerald Lab filter
- Funke Lab (1) Apply Funke Lab filter
- Gonen Lab (2) Apply Gonen Lab filter
- Grigorieff Lab (4) Apply Grigorieff Lab filter
- Harris Lab (4) Apply Harris Lab filter
- Heberlein Lab (2) Apply Heberlein Lab filter
- Hermundstad Lab (1) Apply Hermundstad Lab filter
- Hess Lab (5) Apply Hess Lab filter
- Jayaraman Lab (4) Apply Jayaraman Lab filter
- Ji Lab (1) Apply Ji Lab filter
- Johnson Lab (1) Apply Johnson Lab filter
- Keleman Lab (2) Apply Keleman Lab filter
- Keller Lab (6) Apply Keller Lab filter
- Koay Lab (5) Apply Koay Lab filter
- Lavis Lab (6) Apply Lavis Lab filter
- Lee (Albert) Lab (1) Apply Lee (Albert) Lab filter
- Li Lab (2) Apply Li Lab filter
- Lippincott-Schwartz Lab (12) Apply Lippincott-Schwartz Lab filter
- Liu (Zhe) Lab (7) Apply Liu (Zhe) Lab filter
- Looger Lab (15) Apply Looger Lab filter
- O'Shea Lab (1) Apply O'Shea Lab filter
- Otopalik Lab (3) Apply Otopalik Lab filter
- Pachitariu Lab (4) Apply Pachitariu Lab filter
- Pavlopoulos Lab (1) Apply Pavlopoulos Lab filter
- Pedram Lab (3) Apply Pedram Lab filter
- Podgorski Lab (4) Apply Podgorski Lab filter
- Reiser Lab (2) Apply Reiser Lab filter
- Romani Lab (3) Apply Romani Lab filter
- Rubin Lab (6) Apply Rubin Lab filter
- Saalfeld Lab (3) Apply Saalfeld Lab filter
- Satou Lab (1) Apply Satou Lab filter
- Scheffer Lab (2) Apply Scheffer Lab filter
- Schreiter Lab (4) Apply Schreiter Lab filter
- Simpson Lab (1) Apply Simpson Lab filter
- Singer Lab (4) Apply Singer Lab filter
- Spruston Lab (6) Apply Spruston Lab filter
- Stern Lab (5) Apply Stern Lab filter
- Sternson Lab (2) Apply Sternson Lab filter
- Stringer Lab (4) Apply Stringer Lab filter
- Svoboda Lab (14) Apply Svoboda Lab filter
- Tebo Lab (2) Apply Tebo Lab filter
- Tillberg Lab (2) Apply Tillberg Lab filter
- Truman Lab (4) Apply Truman Lab filter
- Turaga Lab (2) Apply Turaga Lab filter
- Turner Lab (2) Apply Turner Lab filter
- Wang (Shaohe) Lab (4) Apply Wang (Shaohe) Lab filter
- Zlatic Lab (1) Apply Zlatic Lab filter
Associated Project Team
Publication Date
- December 2019 (9) Apply December 2019 filter
- November 2019 (12) Apply November 2019 filter
- October 2019 (20) Apply October 2019 filter
- September 2019 (15) Apply September 2019 filter
- August 2019 (15) Apply August 2019 filter
- July 2019 (15) Apply July 2019 filter
- June 2019 (22) Apply June 2019 filter
- May 2019 (13) Apply May 2019 filter
- April 2019 (18) Apply April 2019 filter
- March 2019 (21) Apply March 2019 filter
- February 2019 (20) Apply February 2019 filter
- January 2019 (22) Apply January 2019 filter
- Remove 2019 filter 2019
Type of Publication
202 Publications
Showing 71-80 of 202 resultsAscidian species of the Phallusia and Ciona genera are distantly related, their last common ancestor dating several hundred million years ago. Although their genome sequences have extensively diverged since this radiation, Phallusia and Ciona species share almost identical early morphogenesis and stereotyped cell lineages. Here, we explored the evolution of transcriptional control between P. mammillata and C. robusta. We combined genome-wide mapping of open chromatin regions in both species with a comparative analysis of the regulatory sequences of a test set of 10 pairs of orthologous early regulatory genes with conserved expression patterns. We find that ascidian chromatin accessibility landscapes obey similar rules as in other metazoa. Open-chromatin regions are short, highly conserved within each genus and cluster around regulatory genes. The dynamics of chromatin accessibility and closest-gene expression are strongly correlated during early embryogenesis. Open-chromatin regions are highly enriched in cis-regulatory elements: 73% of 49 open chromatin regions around our test genes behaved as either distal enhancers or proximal enhancer/promoters following electroporation in Phallusia eggs. Analysis of this datasets suggests a pervasive use in ascidians of "shadow" enhancers with partially overlapping activities. Cross-species electroporations point to a deep conservation of both the trans-regulatory logic between these distantly-related ascidians and the cis-regulatory activities of individual enhancers. Finally, we found that the relative order and approximate distance to the transcription start site of open chromatin regions can be conserved between Ciona and Phallusia species despite extensive sequence divergence, a property that can be used to identify orthologous enhancers, whose regulatory activity can partially diverge.
Genetically wired neural mechanisms inhibit mating between species because even naive animals rarely mate with other species. These mechanisms can evolve through changes in expression or function of key genes in sensory pathways or central circuits. Gr32a is a gustatory chemoreceptor that, in D. melanogaster, is essential to inhibit interspecies courtship and sense quinine. Similar to D. melanogaster, we find that D. simulans Gr32a is expressed in foreleg tarsi, sensorimotor appendages that inhibit interspecies courtship, and it is required to sense quinine. Nevertheless, Gr32a is not required to inhibit interspecies mating by D. simulans males. However, and similar to its function in D. melanogaster, Ppk25, a member of the Pickpocket family, promotes conspecific courtship in D. simulans. Together, we have identified distinct evolutionary mechanisms underlying chemosensory control of taste and courtship in closely related Drosophila species.
Expansion microscopy (ExM) is a physical form of magnification that increases the effective resolving power of any microscope. Here, we describe the fundamental principles of ExM, as well as how recently developed ExM variants build upon and apply those principles. We examine applications of ExM in cell and developmental biology for the study of nanoscale structures as well as ExM's potential for scalable mapping of nanoscale structures across large sample volumes. Finally, we explore how the unique anchoring and hydrogel embedding properties enable postexpansion molecular interrogation in a purified chemical environment. ExM promises to play an important role complementary to emerging live-cell imaging techniques, because of its relative ease of adoption and modification and its compatibility with tissue specimens up to at least 200 μm thick. Expected final online publication date for the , Volume 35 is October 7, 2019. Please see http://www.annualreviews.org/page/journal/pubdates for revised estimates.
Summary This review describes how direct visualization of the dynamic interactions of cells with different extracellular matrix microenvironments can provide novel insights into complex biological processes. Recent studies have moved characterization of cell migration and invasion from classical 2D culture systems into 1D and 3D model systems, revealing multiple differences in mechanisms of cell adhesion, migration and signalling—even though cells in 3D can still display prominent focal adhesions. Myosin II restrains cell migration speed in 2D culture but is often essential for effective 3D migration. 3D cell migration modes can switch between lamellipodial, lobopodial and/or amoeboid depending on the local matrix environment. For example, “nuclear piston” migration can be switched off by local proteolysis, and proteolytic invadopodia can be induced by a high density of fibrillar matrix. Particularly, complex remodelling of both extracellular matrix and tissues occurs during morphogenesis. Extracellular matrix supports self-assembly of embryonic tissues, but it must also be locally actively remodelled. For example, surprisingly focal remodelling of the basement membrane occurs during branching morphogenesis—numerous tiny perforations generated by proteolysis and actomyosin contractility produce a microscopically porous, flexible basement membrane meshwork for tissue expansion. Cells extend highly active blebs or protrusions towards the surrounding mesenchyme through these perforations. Concurrently, the entire basement membrane undergoes translocation in a direction opposite to bud expansion. Underlying this slowly moving 2D basement membrane translocation are highly dynamic individual cell movements. We conclude this review by describing a variety of exciting research opportunities for discovering novel insights into cell-matrix interactions.
Understanding genetic and cellular bases of adult form remains a fundamental goal at the intersection of developmental and evolutionary biology. The skin pigment cells of vertebrates, derived from embryonic neural crest, are a useful system for elucidating mechanisms of fate specification, pattern formation, and how particular phenotypes impact organismal behavior and ecology. In a survey of fishes, including the zebrafish , we identified two populations of white pigment cells-leucophores-one of which arises by transdifferentiation of adult melanophores and another of which develops from a yellow-orange xanthophore or xanthophore-like progenitor. Single-cell transcriptomic, mutational, chemical, and ultrastructural analyses of zebrafish leucophores revealed cell-type-specific chemical compositions, organelle configurations, and genetic requirements. At the organismal level, we identified distinct physiological responses of leucophores during environmental background matching, and we showed that leucophore complement influences behavior. Together, our studies reveal independently arisen pigment cell types and mechanisms of fate acquisition in zebrafish and illustrate how concerted analyses across hierarchical levels can provide insights into phenotypes and their evolution.
Most sensory systems are organized into parallel neuronal pathways that process distinct aspects of incoming stimuli. In the insect olfactory system, second order projection neurons target both the mushroom body, required for learning, and the lateral horn (LH), proposed to mediate innate olfactory behavior. Mushroom body neurons form a sparse olfactory population code, which is not stereotyped across animals. In contrast, odor coding in the LH remains poorly understood. We combine genetic driver lines, anatomical and functional criteria to show that the LH has ~1400 neurons and >165 cell types. Genetically labeled LHNs have stereotyped odor responses across animals and on average respond to three times more odors than single projection neurons. LHNs are better odor categorizers than projection neurons, likely due to stereotyped pooling of related inputs. Our results reveal some of the principles by which a higher processing area can extract innate behavioral significance from sensory stimuli.
The active properties of dendrites can support local nonlinear operations, but previous imaging and electrophysiological measurements have produced conflicting views regarding the prevalence and selectivity of local nonlinearities in vivo. We imaged calcium signals in pyramidal cell dendrites in the motor cortex of mice performing a tactile decision task. A custom microscope allowed us to image the soma and up to 300 μm of contiguous dendrite at 15 Hz, while resolving individual spines. New analysis methods were used to estimate the frequency and spatial scales of activity in dendritic branches and spines. The majority of dendritic calcium transients were coincident with global events. However, task-associated calcium signals in dendrites and spines were compartmentalized by dendritic branching and clustered within branches over approximately 10 μm. Diverse behavior-related signals were intermingled and distributed throughout the dendritic arbor, potentially supporting a large learning capacity in individual neurons.
Plexins exhibit multitudinous, evolutionarily conserved functions in neural development. How Plexins employ their diverse structural motifs in vivo to perform distinct roles is unclear. We previously reported that Plexin B (PlexB) controls multiple steps during the assembly of the olfactory circuit (Li et al., 2018b). Here, we systematically mutagenized structural motifs of PlexB and examined the function of these variants in these multiple steps: axon fasciculation, trajectory choice, and synaptic partner selection. We found that the extracellular Sema domain is essential for all three steps, the catalytic site of the intracellular RapGAP is engaged in none, and the intracellular GTPase-binding motifs are essential for trajectory choice and synaptic partner selection, but are dispensable for fasciculation. Moreover, extracellular PlexB cleavage serves as a regulatory mechanism of PlexB signaling. Thus, the divergent roles of PlexB motifs in distinct steps of neural development contribute to its functional versatility in neural circuit assembly.
Focused Ion Beam Scanning Electron Microscopy (FIB-SEM) generates 3D datasets optimally suited for segmentation of cell ultrastructure and automated connectome tracing but is limited to small fields of view and is therefore incompatible with the new generation of ultrafast multibeam SEMs. In contrast, section-based techniques are multibeam-compatible but are limited in z-resolution making automatic segmentation of cellular ultrastructure difficult. Here we demonstrate a novel 3D electron microscopy technique, Gas Cluster Ion Beam SEM (GCIB-SEM), in which top-down, wide-area ion milling is performed on a series of thick sections, acquiring < 10 nm isotropic datasets of each which are then stitched together to span the full sectioned volume. Based on our results, incorporating GCIB-SEM into existing single beam and multibeam SEM workflows should be straightforward and should dramatically increase reliability while simultaneously improving z-resolution by a factor of 3 or more.
View Publication Page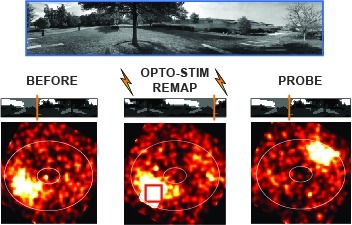
Many animals rely on an internal heading representation when navigating in varied environments. How this representation is linked to the sensory cues that define different surroundings is unclear. In the fly brain, heading is represented by 'compass' neurons that innervate a ring-shaped structure known as the ellipsoid body. Each compass neuron receives inputs from 'ring' neurons that are selective for particular visual features; this combination provides an ideal substrate for the extraction of directional information from a visual scene. Here we combine two-photon calcium imaging and optogenetics in tethered flying flies with circuit modelling, and show how the correlated activity of compass and visual neurons drives plasticity, which flexibly transforms two-dimensional visual cues into a stable heading representation. We also describe how this plasticity enables the fly to convert a partial heading representation, established from orienting within part of a novel setting, into a complete heading representation. Our results provide mechanistic insight into the memory-related computations that are essential for flexible navigation in varied surroundings.