Filter
Associated Lab
- Ahrens Lab (1) Apply Ahrens Lab filter
- Aso Lab (1) Apply Aso Lab filter
- Baker Lab (3) Apply Baker Lab filter
- Betzig Lab (7) Apply Betzig Lab filter
- Bock Lab (2) Apply Bock Lab filter
- Branson Lab (1) Apply Branson Lab filter
- Cardona Lab (1) Apply Cardona Lab filter
- Cui Lab (3) Apply Cui Lab filter
- Dickson Lab (2) Apply Dickson Lab filter
- Druckmann Lab (1) Apply Druckmann Lab filter
- Dudman Lab (1) Apply Dudman Lab filter
- Eddy/Rivas Lab (5) Apply Eddy/Rivas Lab filter
- Fetter Lab (4) Apply Fetter Lab filter
- Fitzgerald Lab (1) Apply Fitzgerald Lab filter
- Gonen Lab (5) Apply Gonen Lab filter
- Grigorieff Lab (6) Apply Grigorieff Lab filter
- Heberlein Lab (12) Apply Heberlein Lab filter
- Hermundstad Lab (1) Apply Hermundstad Lab filter
- Hess Lab (2) Apply Hess Lab filter
- Jayaraman Lab (2) Apply Jayaraman Lab filter
- Ji Lab (2) Apply Ji Lab filter
- Kainmueller Lab (1) Apply Kainmueller Lab filter
- Keller Lab (4) Apply Keller Lab filter
- Lavis Lab (5) Apply Lavis Lab filter
- Lee (Albert) Lab (1) Apply Lee (Albert) Lab filter
- Leonardo Lab (2) Apply Leonardo Lab filter
- Lippincott-Schwartz Lab (18) Apply Lippincott-Schwartz Lab filter
- Liu (Zhe) Lab (1) Apply Liu (Zhe) Lab filter
- Looger Lab (7) Apply Looger Lab filter
- Magee Lab (1) Apply Magee Lab filter
- Menon Lab (3) Apply Menon Lab filter
- Murphy Lab (1) Apply Murphy Lab filter
- Pastalkova Lab (1) Apply Pastalkova Lab filter
- Pavlopoulos Lab (2) Apply Pavlopoulos Lab filter
- Reiser Lab (2) Apply Reiser Lab filter
- Riddiford Lab (1) Apply Riddiford Lab filter
- Romani Lab (1) Apply Romani Lab filter
- Rubin Lab (4) Apply Rubin Lab filter
- Satou Lab (3) Apply Satou Lab filter
- Scheffer Lab (2) Apply Scheffer Lab filter
- Schreiter Lab (3) Apply Schreiter Lab filter
- Sgro Lab (2) Apply Sgro Lab filter
- Simpson Lab (3) Apply Simpson Lab filter
- Singer Lab (10) Apply Singer Lab filter
- Spruston Lab (1) Apply Spruston Lab filter
- Stern Lab (4) Apply Stern Lab filter
- Sternson Lab (6) Apply Sternson Lab filter
- Svoboda Lab (7) Apply Svoboda Lab filter
- Tjian Lab (4) Apply Tjian Lab filter
- Truman Lab (1) Apply Truman Lab filter
- Turaga Lab (1) Apply Turaga Lab filter
- Turner Lab (2) Apply Turner Lab filter
- Zlatic Lab (1) Apply Zlatic Lab filter
- Zuker Lab (3) Apply Zuker Lab filter
Associated Project Team
Publication Date
- December 2011 (22) Apply December 2011 filter
- November 2011 (15) Apply November 2011 filter
- October 2011 (14) Apply October 2011 filter
- September 2011 (17) Apply September 2011 filter
- August 2011 (14) Apply August 2011 filter
- July 2011 (10) Apply July 2011 filter
- June 2011 (17) Apply June 2011 filter
- May 2011 (13) Apply May 2011 filter
- April 2011 (11) Apply April 2011 filter
- March 2011 (14) Apply March 2011 filter
- February 2011 (16) Apply February 2011 filter
- January 2011 (27) Apply January 2011 filter
- Remove 2011 filter 2011
Type of Publication
190 Publications
Showing 101-110 of 190 resultsA reduced sensitivity to the sedating effects of alcohol is a characteristic associated with alcohol use disorders (AUDs). A genetic screen for ethanol sedation mutants in Drosophila identified arouser (aru), which functions in developing neurons to reduce ethanol sensitivity. Genetic evidence suggests that aru regulates ethanol sensitivity through its activation by Egfr/Erk signaling and its inhibition by PI3K/Akt signaling. The aru mutant also has an increased number of synaptic terminals in the larva and adult fly. Both the increased ethanol sensitivity and synapse number of the aru mutant are restored upon adult social isolation, suggesting a causal relationship between synapse number and ethanol sensitivity. We thus show that a developmental abnormality affecting synapse number and ethanol sensitivity is not permanent and can be reversed by manipulating the environment of the adult fly.
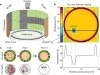
The ability of insects to learn and navigate to specific locations in the environment has fascinated naturalists for decades. The impressive navigational abilities of ants, bees, wasps and other insects demonstrate that insects are capable of visual place learning, but little is known about the underlying neural circuits that mediate these behaviours. Drosophila melanogaster (common fruit fly) is a powerful model organism for dissecting the neural circuitry underlying complex behaviours, from sensory perception to learning and memory. Drosophila can identify and remember visual features such as size, colour and contour orientation. However, the extent to which they use vision to recall specific locations remains unclear. Here we describe a visual place learning platform and demonstrate that Drosophila are capable of forming and retaining visual place memories to guide selective navigation. By targeted genetic silencing of small subsets of cells in the Drosophila brain, we show that neurons in the ellipsoid body, but not in the mushroom bodies, are necessary for visual place learning. Together, these studies reveal distinct neuroanatomical substrates for spatial versus non-spatial learning, and establish Drosophila as a powerful model for the study of spatial memories.

When the contrast of an image flickers as it moves, humans perceive an illusory reversal in the direction of motion. This classic illusion, called reverse-phi motion, has been well-characterized using psychophysics, and several models have been proposed to account for its effects. Here, we show that Drosophila melanogaster also respond behaviorally to the reverse-phi illusion and that the illusion is present in dendritic calcium signals of motion-sensitive neurons in the fly lobula plate. These results closely match the predictions of the predominant model of fly motion detection. However, high flicker rates cause an inversion of the reverse-phi behavioral response that is also present in calcium signals of lobula plate tangential cell dendrites but not predicted by the model. The fly’s behavioral and neural responses to the reverse-phi illusion reveal unexpected interactions between motion and flicker signals in the fly visual system and suggest that a similar correlation-based mechanism underlies visual motion detection across the animal kingdom.
We designed a real-time computer vision system, the Multi-Worm Tracker (MWT), which can simultaneously quantify the behavior of dozens of Caenorhabditis elegans on a Petri plate at video rates. We examined three traditional behavioral paradigms using this system: spontaneous movement on food, where the behavior changes over tens of minutes; chemotaxis, where turning events must be detected accurately to determine strategy; and habituation of response to tap, where the response is stochastic and changes over time. In each case, manual analysis or automated single-worm tracking would be tedious and time-consuming, but the MWT system allowed rapid quantification of behavior with minimal human effort. Thus, this system will enable large-scale forward and reverse genetic screens for complex behaviors.
We analyze the responses of human observers to an ensemble of monomolecular odorants. Each odorant is characterized by a set of 146 perceptual descriptors obtained from a database of odor character profiles. Each odorant is therefore represented by a point in a highly multidimensional sensory space. In this work we study the arrangement of odorants in this perceptual space. We argue that odorants densely sample a two-dimensional curved surface embedded in the multidimensional sensory space. This surface can account for more than half of the variance of the perceptual data. We also show that only 12% of experimental variance cannot be explained by curved surfaces of substantially small dimensionality (<10). We suggest that these curved manifolds represent the relevant spaces sampled by the human olfactory system, thereby providing surrogates for olfactory sensory space. For the case of 2D approximation, we relate the two parameters on the curved surface to the physico-chemical parameters of odorant molecules. We show that one of the dimensions is related to eigenvalues of molecules’ connectivity matrix, while the other is correlated with measures of molecules’ polarity. We discuss the behavioral significance of these findings.
Genetic model organisms such as Drosophila, C. elegans and the mouse provide formidable tools for studying mechanisms of development, physiology and behaviour. Established models alone, however, allow us to survey only a tiny fraction of the morphological and functional diversity present in the animal kingdom. Here, we present iTRAC, a versatile gene-trapping approach that combines the implementation of unbiased genetic screens with the generation of sophisticated genetic tools both in established and emerging model organisms. The approach utilises an exon-trapping transposon vector that carries an integrase docking site, allowing the targeted integration of new constructs into trapped loci. We provide proof of principle for iTRAC in the emerging model crustacean Parhyale hawaiensis: we generate traps that allow specific developmental and physiological processes to be visualised in unparalleled detail, we show that trapped genes can be easily cloned from an unsequenced genome, and we demonstrate targeting of new constructs into a trapped locus. Using this approach, gene traps can serve as platforms for generating diverse reporters, drivers for tissue-specific expression, gene knockdown and other genetic tools not yet imagined.
Analyzing Drosophila melanogaster neural expression patterns in thousands of three-dimensional image stacks of individual brains requires registering them into a canonical framework based on a fiducial reference of neuropil morphology. Given a target brain labeled with predefined landmarks, the BrainAligner program automatically finds the corresponding landmarks in a subject brain and maps it to the coordinate system of the target brain via a deformable warp. Using a neuropil marker (the antibody nc82) as a reference of the brain morphology and a target brain that is itself a statistical average of data for 295 brains, we achieved a registration accuracy of 2 μm on average, permitting assessment of stereotypy, potential connectivity and functional mapping of the adult fruit fly brain. We used BrainAligner to generate an image pattern atlas of 2954 registered brains containing 470 different expression patterns that cover all the major compartments of the fly brain.
Gamma-aminobutyric acid (GABA) is the major inhibitory neurotransmitter in the mammalian brain. Once released, it is removed from the extracellular space by cellular uptake catalyzed by GABA transporter proteins. Four GABA transporters (GAT1, GAT2, GAT3 and BGT1) have been identified. Inhibition of the GAT1 by the clinically available anti-epileptic drug tiagabine has been an effective strategy for the treatment of some patients with partial seizures. Recently, the investigational drug EF1502, which inhibits both GAT1 and BGT1, was found to exert an anti-convulsant action synergistic to that of tiagabine, supposedly due to inhibition of BGT1. The present study addresses the role of BGT1 in seizure control and the effect of EF1502 by developing and exploring a new mouse line lacking exons 3-5 of the BGT1 (slc6a12) gene. The deletion of this sequence abolishes the expression of BGT1 mRNA. However, homozygous BGT1-deficient mice have normal development and show seizure susceptibility indistinguishable from that in wild-type mice in a variety of seizure threshold models including: corneal kindling, the minimal clonic and minimal tonic extension seizure threshold tests, the 6Hz seizure threshold test, and the i.v. pentylenetetrazol threshold test. We confirm that BGT1 mRNA is present in the brain, but find that the levels are several hundred times lower than those of GAT1 mRNA; possibly explaining the apparent lack of phenotype. In conclusion, the present results do not support a role for BGT1 in the control of seizure susceptibility and cannot provide a mechanistic understanding of the synergism that has been previously reported with tiagabine and EF1502.
The formation of amyloid fibrils, protofibrils and oligomers from the β-amyloid (Aβ) peptide represents a hallmark of Alzheimer’s disease. Aβ-peptide-derived assemblies might be crucial for disease onset, but determining their atomic structures has proven to be a major challenge. Progress over the past 5 years has yielded substantial new data obtained with improved methodologies including electron cryo-microscopy and NMR. It is now possible to resolve the global fibril topology and the cross-β sheet organization within protofilaments, and to identify residues that are crucial for stabilizing secondary structural elements and peptide conformations within specific assemblies. These data have significantly enhanced our understanding of the mechanism of Aβ aggregation and have illuminated the possible relevance of specific conformers for neurodegenerative pathologies.
Gap junctions play an important role in the regulation of neuronal metabolism and homeostasis by serving as connections that enable small molecules to pass between cells and synchronize activity between cells. Although recent studies have linked gap junctions to memory formation, it remains unclear how they contribute to this process. Gap junctions are hexameric hemichannels formed from the connexin and pannexin gene families in chordates and the innexin (inx) gene family in invertebrates. Here we show that two modulatory neurons, the anterior paired lateral (APL) neuron and the dorsal paired medial (DPM) neuron, form heterotypic gap junctions within the mushroom body (MB), a learning and memory center in the Drosophila brain. Using RNA interference-mediated knockdowns of inx7 and inx6 in the APL and DPM neurons, respectively, we found that flies showed normal olfactory associative learning and intact anesthesia-resistant memory (ARM) but failed to form anesthesia-sensitive memory (ASM). Our results reveal that the heterotypic gap junctions between the APL and DPM neurons are an essential part of the MB circuitry for memory formation, potentially constituting a recurrent neural network to stabilize ASM.