Filter
Associated Lab
- Card Lab (3) Apply Card Lab filter
- Dickson Lab (5) Apply Dickson Lab filter
- Funke Lab (2) Apply Funke Lab filter
- Lavis Lab (2) Apply Lavis Lab filter
- Reiser Lab (1) Apply Reiser Lab filter
- Rubin Lab (1) Apply Rubin Lab filter
- Singer Lab (2) Apply Singer Lab filter
- Remove Stern Lab filter Stern Lab
- Tillberg Lab (2) Apply Tillberg Lab filter
- Truman Lab (4) Apply Truman Lab filter
Associated Project Team
Publication Date
- 2025 (4) Apply 2025 filter
- 2024 (9) Apply 2024 filter
- 2023 (2) Apply 2023 filter
- 2022 (8) Apply 2022 filter
- 2021 (5) Apply 2021 filter
- 2020 (4) Apply 2020 filter
- 2019 (5) Apply 2019 filter
- 2018 (6) Apply 2018 filter
- 2017 (8) Apply 2017 filter
- 2016 (7) Apply 2016 filter
- 2015 (4) Apply 2015 filter
- 2014 (6) Apply 2014 filter
- 2013 (6) Apply 2013 filter
- 2012 (5) Apply 2012 filter
- 2011 (4) Apply 2011 filter
- 2010 (8) Apply 2010 filter
- 2009 (5) Apply 2009 filter
- 2008 (4) Apply 2008 filter
- 2007 (9) Apply 2007 filter
- 2006 (6) Apply 2006 filter
- 2005 (6) Apply 2005 filter
- 2004 (3) Apply 2004 filter
- 2003 (8) Apply 2003 filter
- 2001 (1) Apply 2001 filter
- 2000 (4) Apply 2000 filter
- 1999 (2) Apply 1999 filter
- 1998 (2) Apply 1998 filter
- 1997 (3) Apply 1997 filter
- 1996 (3) Apply 1996 filter
- 1995 (2) Apply 1995 filter
- 1994 (2) Apply 1994 filter
- 1993 (1) Apply 1993 filter
- 1991 (3) Apply 1991 filter
- 1990 (1) Apply 1990 filter
Type of Publication
156 Publications
Showing 131-140 of 156 resultsAphids exhibit divergent modes of embryogenesis during the sexual and asexual phases of the life cycle. To explore how a single genome can give rise to these alternative developmental modes, we have initiated embryological studies of the pea aphid, Acyrthosiphon pisum. Here we present a detailed description of parthenogenetic, viviparous embryonic development in the pea aphid. We compare and contrast development of the parthenogenetic embryo with that of the embryo resulting from sexual reproduction. The primary difference between the embryos is the scale on which development occurs: early parthenogenetic development occurs in a volume approximately three orders of magnitude smaller than the sexual egg, largely because of the apparent absence of yolk in the parthenogenetic egg. This results in a drastically different duration of syncytial energid cleavage and, presumably, patterning processes in the two embryos must act at scales that differ by orders of magnitude. The eggs also develop on time scales that differ approximately by an order of magnitude and the timing of the embryonic movements, collectively called blastokinesis, have temporally shifted relative to growth of the embryo. In addition, the endosymbiotic bacteria are transferred from mother to embryo in different ways in the two embryos. Finally, the function of the serosa has diverged greatly in the two embryos: in the sexual egg the serosa deposits a thick cuticle that protects the egg, whereas the serosa of the parthenogenetic embryo is greatly reduced and its function is unclear. The pea aphid is a useful model system for examining how a single genome has evolved to allow divergent modes of development.
Mutualism with ants is suspected to be a highly labile trait within homopteran evolution. We used molecular phylogenetic evidence to test whether the mutualism has multiple origins within a single aphid genus. We constructed a molecular phylogeny of 15 Chaitophorus Koch (Hemiptera: Aphidoidea) species, using mitochondrial cytochrome oxidase I and II sequences. Ant tending evolved, or was lost, at least five times during Chaitophorus evolution. Parametric bootstrapping rejected the hypothesis of a single origin of ant tending in this genus. Further, the Chaitophorus made at least two host genus switches from poplars (Populus) to willow (Salix), and four switches in feeding position, from leaf feeding to stem feeding or vice versa. This is the first phylogenetic confirmation that ant tending is an evolutionarily labile trait in aphids.
Separate recent studies have revealed the physiological changes underlying the evolution of body size in an insect and advanced our understanding of the genetics of insect growth. These studies highlight the gulf between physiological and genetic studies of growth control and the exciting opportunities for unification of these fields.
One of the oldest problems in evolutionary biology remains largely unsolved. Which mutations generate evolutionarily relevant phenotypic variation? What kinds of molecular changes do they entail? What are the phenotypic magnitudes, frequencies of origin, and pleiotropic effects of such mutations? How is the genome constructed to allow the observed abundance of phenotypic diversity? Historically, the neo-Darwinian synthesizers stressed the predominance of micromutations in evolution, whereas others noted the similarities between some dramatic mutations and evolutionary transitions to argue for macromutationism. Arguments on both sides have been biased by misconceptions of the developmental effects of mutations. For example, the traditional view that mutations of important developmental genes always have large pleiotropic effects can now be seen to be a conclusion drawn from observations of a small class of mutations with dramatic effects. It is possible that some mutations, for example, those in cis-regulatory DNA, have few or no pleiotropic effects and may be the predominant source of morphological evolution. In contrast, mutations causing dramatic phenotypic effects, although superficially similar to hypothesized evolutionary transitions, are unlikely to fairly represent the true path of evolution. Recent developmental studies of gene function provide a new way of conceptualizing and studying variation that contrasts with the traditional genetic view that was incorporated into neo-Darwinian theory and population genetics. This new approach in developmental biology is as important for microevolutionary studies as the actual results from recent evolutionary developmental studies. In particular, this approach will assist in the task of identifying the specific mutations generating phenotypic variation and elucidating how they alter gene function. These data will provide the current missing link between molecular and phenotypic variation in natural populations.
We report an extreme morphological difference between Drosophila sechellia and related species of the pattern of hairs on first-instar larvae. On the dorsum of most species, the posterior region of the anterior compartment of most segments is covered by a carpet of fine hairs. In D. sechellia, these hairs have been lost and replaced with naked cuticle. Genetic mapping experiments and interspecific complementation tests indicate that this difference is caused, in its entirety, by evolution at the ovo/shaven-baby locus. The pattern of expression of the ovo/shaven-baby transcript is correlated with this morphological change. The altered dorsal cuticle pattern is probably caused by evolution of the cis-regulatory region of ovo/shaven-baby in the D. sechellia lineage.
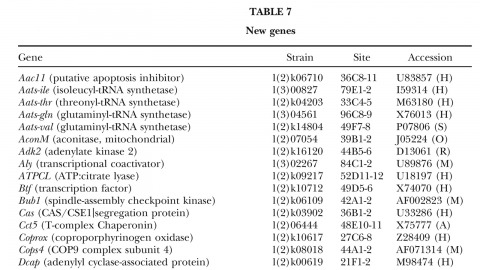
A fundamental goal of genetics and functional genomics is to identify and mutate every gene in model organisms such as Drosophila melanogaster. The Berkeley Drosophila Genome Project (BDGP) gene disruption project generates single P-element insertion strains that each mutate unique genomic open reading frames. Such strains strongly facilitate further genetic and molecular studies of the disrupted loci, but it has remained unclear if P elements can be used to mutate all Drosophila genes. We now report that the primary collection has grown to contain 1045 strains that disrupt more than 25% of the estimated 3600 Drosophila genes that are essential for adult viability. Of these P insertions, 67% have been verified by genetic tests to cause the associated recessive mutant phenotypes, and the validity of most of the remaining lines is predicted on statistical grounds. Sequences flanking >920 insertions have been determined to exactly position them in the genome and to identify 376 potentially affected transcripts from collections of EST sequences. Strains in the BDGP collection are available from the Bloomington Stock Center and have already assisted the research community in characterizing >250 Drosophila genes. The likely identity of 131 additional genes in the collection is reported here. Our results show that Drosophila genes have a wide range of sensitivity to inactivation by P elements, and provide a rationale for greatly expanding the BDGP primary collection based entirely on insertion site sequencing. We predict that this approach can bring >85% of all Drosophila open reading frames under experimental control.
Within all species of animals, the size of each organ bears a specific relationship to overall body size. These patterns of organ size relative to total body size are called static allometry and have enchanted biologists for centuries, yet the mechanisms generating these patterns have attracted little experimental study. We review recent and older work on holometabolous insect development that sheds light on these mechanisms. In insects, static allometry can be divided into at least two processes: (1) the autonomous specification of organ identity, perhaps including the approximate size of the organ, and (2) the determination of the final size of organs based on total body size. We present three models to explain the second process: (1) all organs autonomously absorb nutrients and grow at organ-specific rates, (2) a centralized system measures a close correlate of total body size and distributes this information to all organs, and (3) autonomous organ growth is combined with feedback between growing organs to modulate final sizes. We provide evidence supporting models 2 and 3 and also suggest that hormones are the messengers of size information. Advances in our understanding of the mechanisms of allometry will come through the integrated study of whole tissues using techniques from development, genetics, endocrinology and population biology.
The mechanisms underlying the evolution of morphology are poorly understood. Distantly related taxa sometimes exhibit correlations between morphological differences and patterns of gene expression, but such comparisons cannot establish how mechanisms evolve to generate diverse morphologies. Answers to these questions require resolution of the nature of developmental evolution within and between closely related species. Here I show how the detailed regulation of the Hox gene Ultrabithorax patterns trichomes on the posterior femur of the second leg in Drosophila melanogaster, and that evolution of Ultrabithorax has contributed to divergence of this feature among closely related species. The cis-regulatory regions of Ultrabithorax, and not the protein itself, appear to have evolved. This study provides experimental evidence that cis-regulatory evolution is one way in which conserved proteins have promoted morphological diversity.