Filter
Associated Lab
- Aguilera Castrejon Lab (15) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (11) Apply Ahrens Lab filter
- Baker Lab (19) Apply Baker Lab filter
- Betzig Lab (12) Apply Betzig Lab filter
- Beyene Lab (5) Apply Beyene Lab filter
- Bock Lab (3) Apply Bock Lab filter
- Branson Lab (3) Apply Branson Lab filter
- Card Lab (6) Apply Card Lab filter
- Cardona Lab (19) Apply Cardona Lab filter
- Chklovskii Lab (3) Apply Chklovskii Lab filter
- Clapham Lab (1) Apply Clapham Lab filter
- Darshan Lab (4) Apply Darshan Lab filter
- Dennis Lab (1) Apply Dennis Lab filter
- Dickson Lab (14) Apply Dickson Lab filter
- Druckmann Lab (4) Apply Druckmann Lab filter
- Dudman Lab (12) Apply Dudman Lab filter
- Egnor Lab (7) Apply Egnor Lab filter
- Espinosa Medina Lab (4) Apply Espinosa Medina Lab filter
- Fetter Lab (10) Apply Fetter Lab filter
- Fitzgerald Lab (13) Apply Fitzgerald Lab filter
- Gonen Lab (32) Apply Gonen Lab filter
- Grigorieff Lab (28) Apply Grigorieff Lab filter
- Harris Lab (10) Apply Harris Lab filter
- Heberlein Lab (81) Apply Heberlein Lab filter
- Hermundstad Lab (4) Apply Hermundstad Lab filter
- Hess Lab (3) Apply Hess Lab filter
- Jayaraman Lab (4) Apply Jayaraman Lab filter
- Johnson Lab (5) Apply Johnson Lab filter
- Kainmueller Lab (19) Apply Kainmueller Lab filter
- Karpova Lab (1) Apply Karpova Lab filter
- Keleman Lab (5) Apply Keleman Lab filter
- Keller Lab (15) Apply Keller Lab filter
- Koay Lab (16) Apply Koay Lab filter
- Lavis Lab (12) Apply Lavis Lab filter
- Lee (Albert) Lab (5) Apply Lee (Albert) Lab filter
- Leonardo Lab (4) Apply Leonardo Lab filter
- Li Lab (24) Apply Li Lab filter
- Lippincott-Schwartz Lab (72) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (5) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (5) Apply Liu (Zhe) Lab filter
- Looger Lab (1) Apply Looger Lab filter
- Magee Lab (18) Apply Magee Lab filter
- Menon Lab (6) Apply Menon Lab filter
- Murphy Lab (7) Apply Murphy Lab filter
- O'Shea Lab (1) Apply O'Shea Lab filter
- Otopalik Lab (12) Apply Otopalik Lab filter
- Pachitariu Lab (12) Apply Pachitariu Lab filter
- Pastalkova Lab (13) Apply Pastalkova Lab filter
- Pavlopoulos Lab (12) Apply Pavlopoulos Lab filter
- Pedram Lab (11) Apply Pedram Lab filter
- Reiser Lab (6) Apply Reiser Lab filter
- Riddiford Lab (24) Apply Riddiford Lab filter
- Romani Lab (12) Apply Romani Lab filter
- Rubin Lab (38) Apply Rubin Lab filter
- Saalfeld Lab (17) Apply Saalfeld Lab filter
- Satou Lab (15) Apply Satou Lab filter
- Schreiter Lab (17) Apply Schreiter Lab filter
- Sgro Lab (20) Apply Sgro Lab filter
- Simpson Lab (5) Apply Simpson Lab filter
- Singer Lab (43) Apply Singer Lab filter
- Spruston Lab (36) Apply Spruston Lab filter
- Stern Lab (83) Apply Stern Lab filter
- Sternson Lab (7) Apply Sternson Lab filter
- Stringer Lab (3) Apply Stringer Lab filter
- Svoboda Lab (4) Apply Svoboda Lab filter
- Tebo Lab (24) Apply Tebo Lab filter
- Tillberg Lab (3) Apply Tillberg Lab filter
- Tjian Lab (47) Apply Tjian Lab filter
- Truman Lab (30) Apply Truman Lab filter
- Turaga Lab (12) Apply Turaga Lab filter
- Turner Lab (11) Apply Turner Lab filter
- Wang (Shaohe) Lab (19) Apply Wang (Shaohe) Lab filter
- Wu Lab (1) Apply Wu Lab filter
- Zlatic Lab (2) Apply Zlatic Lab filter
- Zuker Lab (20) Apply Zuker Lab filter
Associated Project Team
Publication Date
- 2024 (1) Apply 2024 filter
- 2023 (1) Apply 2023 filter
- 2022 (26) Apply 2022 filter
- 2021 (19) Apply 2021 filter
- 2020 (19) Apply 2020 filter
- 2019 (25) Apply 2019 filter
- 2018 (26) Apply 2018 filter
- 2017 (31) Apply 2017 filter
- 2016 (18) Apply 2016 filter
- 2015 (57) Apply 2015 filter
- 2014 (46) Apply 2014 filter
- 2013 (58) Apply 2013 filter
- 2012 (78) Apply 2012 filter
- 2011 (92) Apply 2011 filter
- 2010 (100) Apply 2010 filter
- 2009 (102) Apply 2009 filter
- 2008 (100) Apply 2008 filter
- 2007 (85) Apply 2007 filter
- 2006 (89) Apply 2006 filter
- 2005 (67) Apply 2005 filter
- 2004 (57) Apply 2004 filter
- 2003 (58) Apply 2003 filter
- 2002 (39) Apply 2002 filter
- 2001 (28) Apply 2001 filter
- 2000 (29) Apply 2000 filter
- 1999 (14) Apply 1999 filter
- 1998 (18) Apply 1998 filter
- 1997 (16) Apply 1997 filter
- 1996 (10) Apply 1996 filter
- 1995 (18) Apply 1995 filter
- 1994 (12) Apply 1994 filter
- 1993 (10) Apply 1993 filter
- 1992 (6) Apply 1992 filter
- 1991 (11) Apply 1991 filter
- 1990 (11) Apply 1990 filter
- 1989 (6) Apply 1989 filter
- 1988 (1) Apply 1988 filter
- 1987 (7) Apply 1987 filter
- 1986 (4) Apply 1986 filter
- 1985 (5) Apply 1985 filter
- 1984 (2) Apply 1984 filter
- 1983 (2) Apply 1983 filter
- 1982 (3) Apply 1982 filter
- 1981 (3) Apply 1981 filter
- 1980 (1) Apply 1980 filter
- 1979 (1) Apply 1979 filter
- 1976 (2) Apply 1976 filter
- 1973 (1) Apply 1973 filter
- 1970 (1) Apply 1970 filter
- 1967 (1) Apply 1967 filter
Type of Publication
- Remove Non-Janelia filter Non-Janelia
1417 Publications
Showing 1241-1250 of 1417 resultsI consider a topographic projection between two neuronal layers with different densities of neurons. Given the number of output neurons connected to each input neuron (divergence) and the number of input neurons synapsing on each output neuron (convergence), I determine the widths of axonal and dendritic arbors which minimize the total volume of axons and dendrites. Analytical results for one-dimensional and two-dimensional projections can be summarized qualitatively in the following rule: neurons of the sparser layer should have arbors wider than those of the denser layer. This agrees with the anatomic data for retinal, cerebellar, olfactory bulb, and neocortical neurons the morphology and connectivity of which are known. The rule may be used to infer connectivity of neurons from their morphology.
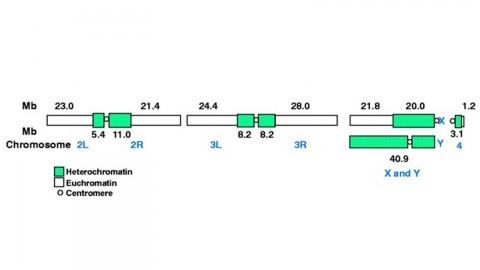
We report on the quality of a whole-genome assembly of Drosophila melanogaster and the nature of the computer algorithms that accomplished it. Three independent external data sources essentially agree with and support the assembly’s sequence and ordering of contigs across the euchromatic portion of the genome. In addition, there are isolated contigs that we believe represent nonrepetitive pockets within the heterochromatin of the centromeres. Comparison with a previously sequenced 2.9- megabase region indicates that sequencing accuracy within nonrepetitive segments is greater than 99. 99% without manual curation. As such, this initial reconstruction of the Drosophila sequence should be of substantial value to the scientific community.

A comparative analysis of the genomes of Drosophila melanogaster, Caenorhabditis elegans, and Saccharomyces cerevisiae-and the proteins they are predicted to encode-was undertaken in the context of cellular, developmental, and evolutionary processes. The nonredundant protein sets of flies and worms are similar in size and are only twice that of yeast, but different gene families are expanded in each genome, and the multidomain proteins and signaling pathways of the fly and worm are far more complex than those of yeast. The fly has orthologs to 177 of the 289 human disease genes examined and provides the foundation for rapid analysis of some of the basic processes involved in human disease.
The fly Drosophila melanogaster is one of the most intensively studied organisms in biology and serves as a model system for the investigation of many developmental and cellular processes common to higher eukaryotes, including humans. We have determined the nucleotide sequence of nearly all of the approximately 120-megabase euchromatic portion of the Drosophila genome using a whole-genome shotgun sequencing strategy supported by extensive clone-based sequence and a high-quality bacterial artificial chromosome physical map. Efforts are under way to close the remaining gaps; however, the sequence is of sufficient accuracy and contiguity to be declared substantially complete and to support an initial analysis of genome structure and preliminary gene annotation and interpretation. The genome encodes approximately 13,600 genes, somewhat fewer than the smaller Caenorhabditis elegans genome, but with comparable functional diversity.
In mammals, taste perception is a major mode of sensory input. We have identified a novel family of 40-80 human and rodent G protein-coupled receptors expressed in subsets of taste receptor cells of the tongue and palate epithelia. These candidate taste receptors (T2Rs) are organized in the genome in clusters and are genetically linked to loci that influence bitter perception in mice and humans. Notably, a single taste receptor cell expresses a large repertoire of T2Rs, suggesting that each cell may be capable of recognizing multiple tastants. T2Rs are exclusively expressed in taste receptor cells that contain the G protein alpha subunit gustducin, implying that they function as gustducin-linked receptors. In the accompanying paper, we demonstrate that T2Rs couple to gustducin in vitro, and respond to bitter tastants in a functional expression assay.
Bitter taste perception provides animals with critical protection against ingestion of poisonous compounds. In the accompanying paper, we report the characterization of a large family of putative mammalian taste receptors (T2Rs). Here we use a heterologous expression system to show that specific T2Rs function as bitter taste receptors. A mouse T2R (mT2R-5) responds to the bitter tastant cycloheximide, and a human and a mouse receptor (hT2R-4 and mT2R-8) responded to denatonium and 6-n-propyl-2-thiouracil. Mice strains deficient in their ability to detect cycloheximide have amino acid substitutions in the mT2R-5 gene; these changes render the receptor significantly less responsive to cycloheximide. We also expressed mT2R-5 in insect cells and demonstrate specific tastant-dependent activation of gustducin, a G protein implicated in bitter signaling. Since a single taste receptor cell expresses a large repertoire of T2Rs, these findings provide a plausible explanation for the uniform bitter taste that is evoked by many structurally unrelated toxic compounds.
The steroid hormone 20-hydroxyecdysone (20E) initiates metamorphosis in insects by signaling through the ecdysone receptor complex, a heterodimer of the ecdysone receptor (EcR) and ultraspiracle (USP). Analysis of usp mutant clones in the wing disc of Drosophila shows that in the absence of USP, early hormone responsive genes such as EcR, DHR3 and E75B fail to up-regulate in response to 20E, but other genes that are normally expressed later, such as (&bgr;)-Ftz-F1 and the Z1 isoform of the Broad-Complex (BRC-Z1), are expressed precociously. Sensory neuron formation and axonal outgrowth, two early metamorphic events, also occur prematurely. In vitro experiments with cultured wing discs showed that BRC-Z1 expression and early metamorphic development are rendered steroid-independent in the usp mutant clones. These results are consistent with a model in which these latter processes are induced by a signal arising during the middle of the last larval stage but suppressed by the unliganded EcR/USP complex. Our observations suggest that silencing by the unliganded EcR/USP receptor and the subsequent release of silencing by moderate steroid levels may play an important role in coordinating early phases of steroid driven development.
Drugs of abuse have a common property in mammals, which is their ability to facilitate the release of the neurotransmitter and neuromodulator dopamine in specific brain regions involved in reward and motivation. This increase in synaptic dopamine levels is believed to act as a positive reinforcer and to mediate some of the acute responses to drugs. The mechanisms by which dopamine regulates acute drug responses and addiction remain unknown.