Filter
Associated Lab
- Aguilera Castrejon Lab (15) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (11) Apply Ahrens Lab filter
- Baker Lab (19) Apply Baker Lab filter
- Betzig Lab (12) Apply Betzig Lab filter
- Beyene Lab (5) Apply Beyene Lab filter
- Bock Lab (3) Apply Bock Lab filter
- Branson Lab (3) Apply Branson Lab filter
- Card Lab (6) Apply Card Lab filter
- Cardona Lab (19) Apply Cardona Lab filter
- Chklovskii Lab (3) Apply Chklovskii Lab filter
- Clapham Lab (1) Apply Clapham Lab filter
- Darshan Lab (4) Apply Darshan Lab filter
- Dennis Lab (1) Apply Dennis Lab filter
- Dickson Lab (14) Apply Dickson Lab filter
- Druckmann Lab (4) Apply Druckmann Lab filter
- Dudman Lab (12) Apply Dudman Lab filter
- Egnor Lab (7) Apply Egnor Lab filter
- Espinosa Medina Lab (4) Apply Espinosa Medina Lab filter
- Fetter Lab (10) Apply Fetter Lab filter
- Fitzgerald Lab (13) Apply Fitzgerald Lab filter
- Gonen Lab (32) Apply Gonen Lab filter
- Grigorieff Lab (28) Apply Grigorieff Lab filter
- Harris Lab (10) Apply Harris Lab filter
- Heberlein Lab (81) Apply Heberlein Lab filter
- Hermundstad Lab (4) Apply Hermundstad Lab filter
- Hess Lab (3) Apply Hess Lab filter
- Jayaraman Lab (4) Apply Jayaraman Lab filter
- Johnson Lab (5) Apply Johnson Lab filter
- Kainmueller Lab (19) Apply Kainmueller Lab filter
- Karpova Lab (1) Apply Karpova Lab filter
- Keleman Lab (5) Apply Keleman Lab filter
- Keller Lab (15) Apply Keller Lab filter
- Koay Lab (16) Apply Koay Lab filter
- Lavis Lab (12) Apply Lavis Lab filter
- Lee (Albert) Lab (5) Apply Lee (Albert) Lab filter
- Leonardo Lab (4) Apply Leonardo Lab filter
- Li Lab (24) Apply Li Lab filter
- Lippincott-Schwartz Lab (72) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (5) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (5) Apply Liu (Zhe) Lab filter
- Looger Lab (1) Apply Looger Lab filter
- Magee Lab (18) Apply Magee Lab filter
- Menon Lab (6) Apply Menon Lab filter
- Murphy Lab (7) Apply Murphy Lab filter
- O'Shea Lab (1) Apply O'Shea Lab filter
- Otopalik Lab (12) Apply Otopalik Lab filter
- Pachitariu Lab (12) Apply Pachitariu Lab filter
- Pastalkova Lab (13) Apply Pastalkova Lab filter
- Pavlopoulos Lab (12) Apply Pavlopoulos Lab filter
- Pedram Lab (11) Apply Pedram Lab filter
- Reiser Lab (6) Apply Reiser Lab filter
- Riddiford Lab (24) Apply Riddiford Lab filter
- Romani Lab (12) Apply Romani Lab filter
- Rubin Lab (38) Apply Rubin Lab filter
- Saalfeld Lab (17) Apply Saalfeld Lab filter
- Satou Lab (15) Apply Satou Lab filter
- Schreiter Lab (17) Apply Schreiter Lab filter
- Sgro Lab (20) Apply Sgro Lab filter
- Simpson Lab (5) Apply Simpson Lab filter
- Singer Lab (43) Apply Singer Lab filter
- Spruston Lab (36) Apply Spruston Lab filter
- Stern Lab (83) Apply Stern Lab filter
- Sternson Lab (7) Apply Sternson Lab filter
- Stringer Lab (3) Apply Stringer Lab filter
- Svoboda Lab (4) Apply Svoboda Lab filter
- Tebo Lab (24) Apply Tebo Lab filter
- Tillberg Lab (3) Apply Tillberg Lab filter
- Tjian Lab (47) Apply Tjian Lab filter
- Truman Lab (30) Apply Truman Lab filter
- Turaga Lab (12) Apply Turaga Lab filter
- Turner Lab (11) Apply Turner Lab filter
- Wang (Shaohe) Lab (19) Apply Wang (Shaohe) Lab filter
- Wu Lab (1) Apply Wu Lab filter
- Zlatic Lab (2) Apply Zlatic Lab filter
- Zuker Lab (20) Apply Zuker Lab filter
Associated Project Team
Publication Date
- 2024 (1) Apply 2024 filter
- 2023 (1) Apply 2023 filter
- 2022 (26) Apply 2022 filter
- 2021 (19) Apply 2021 filter
- 2020 (19) Apply 2020 filter
- 2019 (25) Apply 2019 filter
- 2018 (26) Apply 2018 filter
- 2017 (31) Apply 2017 filter
- 2016 (18) Apply 2016 filter
- 2015 (57) Apply 2015 filter
- 2014 (46) Apply 2014 filter
- 2013 (58) Apply 2013 filter
- 2012 (78) Apply 2012 filter
- 2011 (92) Apply 2011 filter
- 2010 (100) Apply 2010 filter
- 2009 (102) Apply 2009 filter
- 2008 (100) Apply 2008 filter
- 2007 (85) Apply 2007 filter
- 2006 (89) Apply 2006 filter
- 2005 (67) Apply 2005 filter
- 2004 (57) Apply 2004 filter
- 2003 (58) Apply 2003 filter
- 2002 (39) Apply 2002 filter
- 2001 (28) Apply 2001 filter
- 2000 (29) Apply 2000 filter
- 1999 (14) Apply 1999 filter
- 1998 (18) Apply 1998 filter
- 1997 (16) Apply 1997 filter
- 1996 (10) Apply 1996 filter
- 1995 (18) Apply 1995 filter
- 1994 (12) Apply 1994 filter
- 1993 (10) Apply 1993 filter
- 1992 (6) Apply 1992 filter
- 1991 (11) Apply 1991 filter
- 1990 (11) Apply 1990 filter
- 1989 (6) Apply 1989 filter
- 1988 (1) Apply 1988 filter
- 1987 (7) Apply 1987 filter
- 1986 (4) Apply 1986 filter
- 1985 (5) Apply 1985 filter
- 1984 (2) Apply 1984 filter
- 1983 (2) Apply 1983 filter
- 1982 (3) Apply 1982 filter
- 1981 (3) Apply 1981 filter
- 1980 (1) Apply 1980 filter
- 1979 (1) Apply 1979 filter
- 1976 (2) Apply 1976 filter
- 1973 (1) Apply 1973 filter
- 1970 (1) Apply 1970 filter
- 1967 (1) Apply 1967 filter
Type of Publication
- Remove Non-Janelia filter Non-Janelia
1417 Publications
Showing 1311-1320 of 1417 resultsSelection for mutants which release glucose repression of the CYB2 gene was used to identify genes which regulate repression of mitochondrial biogenesis. We have identified two of these as the previously described GRR1/CAT80 and ROX3 genes. Mutations in these genes not only release glucose repression of CYB2 but also generally release respiration of the mutants from glucose repression. In addition, both mutants are partially defective in CYB2 expression when grown on nonfermentable carbon sources, indicating a positive regulatory role as well. ROX3 was cloned by complementation of a glucose-inducible flocculating phenotype of an amber mutant and has been mapped as a new leftmost marker on chromosome 2. The ROX3 mutant has only a modest defect in glucose repression of GAL1 but is substantially compromised in galactose induction of GAL1 expression. This mutant also has increased SUC2 expression on nonrepressing carbon sources. We have also characterized the regulation of CYB2 in strains carrying null mutation in two other glucose repression genes, HXK2 and SSN6, and show that HXK2 is a negative regulator of CYB2, whereas SSN6 appears to be a positive effector of CYB2 expression.
The Drosophila retina is a crystalline array of 800 ommatidia whose organization and assembly suggest polarization of the retinal epithelium along anteroposterior and dorsoventral axes. The retina develops by a stepwise process following the posterior-to-anterior progression of the morphogenetic furrow across the eye disc. Ectopic expression of hedgehog or local removal of patched function generates ectopic furrows that can progress in any direction across the disc leaving in their wake differentiating fields of ectopic ommatidia. We have studied the effect of these ectopic furrows on the polarity of ommatidial assembly and rotation. We find that the anteroposterior asymmetry of ommatidial assembly parallels the progression of ectopic furrows, regardless of their direction. In addition, ommatidia developing behind ectopic furrows rotate coordinately, forming equators in various regions of the disc. Interestingly, the expression of a marker normally restricted to the equator is induced in ectopic ommatidial fields. Ectopic equators are stable as they persist to adulthood, where they can coexist with the normal equator. Our results suggest that ectopic furrows can impart polarity to the disc epithelium, regarding the direction of both assembly and rotation of ommatidia. We propose that these processes are polarized as a consequence of furrow propagation, while more global determinants of dorsoventral and anteroposterior polarity may act less directly by determining the site of furrow initiation.
Humans respond adaptively to uncertainty by escaping or seeking additional information. To foster a comparative study of uncertainty processes, we asked whether humans and a bottlenosed dolphin (Tursiops truncatus) would use similarly a psychophysical uncertain response. Human observers and the dolphin were given 2 primary discrimination responses and a way to escape chosen trials into easier ones. Humans escaped sparingly from the most difficult trials near threshold that left them demonstrably uncertain of the stimulus. The dolphin performed nearly identically. The behavior of both species is considered from the perspectives of signal detection theory and optimality theory, and its appropriate interpretation is discussed. Human and dolphin uncertain responses seem to be interesting cognitive analogs and may depend on cognitive or controlled decisional mechanisms. The capacity to monitor ongoing cognition, and use uncertainty appropriately, would be a valuable adaptation for animal minds. This recommends uncertainty processes as an important but neglected area for future comparative research.
In Drosophila dosage compensation increases the rate of transcription of the male's X chromosome and depends on four autosomal male-specific lethal genes. We have cloned the msl-2 gene and shown that MSL-2 protein is co-localized with the other three MSL proteins at hundreds of sites along the male polytene X chromosome and that this binding requires the other three MSL proteins. msl-2 encodes a protein with a putative DNA-binding domain: the RING finger. MSL-2 protein is not produced in females and sequences in both the 5' and 3' UTRs are important for this sex-specific regulation. Furthermore, msl-2 pre-mRNA is alternatively spliced in a Sex-lethal-dependent fashion in its 5' UTR.
The fragment assembly problem is that of reconstructing a DNA sequence from a collection of randomly sampled fragments. Traditionally, the objective of this problem has been to produce the shortest string that contains all the fragments as substrings, but in the case of repetitive target sequences this objective produces answers that are overcompressed. In this paper, the problem is reformulated as one of finding a maximum-likelihood reconstruction with respect to the two-sided Kolmogorov-Smirnov statistic, and it is argued that this is a better formulation of the problem. Next the fragment assembly problem is recast in graph-theoretic terms as one of finding a noncyclic subgraph with certain properties and the objectives of being shortest or maximally likely are also recast in this framework. Finally, a series of graph reduction transformations are given that dramatically reduce the size of the graph to be explored in practical instances of the problem. This reduction is very important as the underlying problems are NP-hard. In practice, the transformed problems are so small that simple branch-and-bound algorithms successfully solve them, thus permitting auxiliary experimental information to be taken into account in the form of overlap, orientation, and distance constraints.
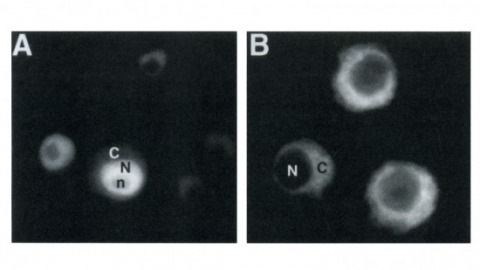
Drosophila yan has been postulated to act as an antagonist of the proneural signal mediated by the sevenless/Ras1/MAPK pathway. We have mutagenized the eight MAPK phosphorylation consensus sites of yan and examined the effects of overexpressing the mutant protein in transgenic flies and transfected S2 cultured cells. Our results suggest that phosphorylation by MAPK affects the stability and subcellular localization of yan, resulting in rapid down-regulation of yan activity. Furthermore, MAPK-mediated down-regulation of yan function appears to be critical for the proper differentiation of both neuronal and nonneuronal tissues throughout development, suggesting that yan is an essential component of a general timing mechanism controlling the competence of a cell to respond to inductive signals.
Most neurons in the CNS have complex, branching dendritic trees, which receive the majority of all synaptic input. As it is difficult to make electrical recordings from dendrites because of their small size, most of what is known about their electrical properties has been inferred from recordings made at the soma. By taking advantage of the higher resolution offered by improved optics, it is now possible to make patch-pipette recordings from the dendrites of neurons in brain slices under visual control. This new technique promises to provide valuable new information concerning dendritic function.
Human mitochondrial transcription factor A (h-mtTFA) is essential for initiation of transcription from the two promoters located in the displacement-loop region of human mitochondrial DNA. This 25 kDa protein contains two tandem, HMG box DNA-binding domains separated by a 27 amino acid residue linker region and followed by a 25 residue carboxyl-terminal tail; both the linker and tail are rich in basic amino acid residues. Mutational analysis of h-mtTFA revealed that the tail region is important for specific DNA recognition and essential for transcriptional activation. The critical role of the human tail in transcription was confirmed by constructing chimeric proteins that exchanged similar regions between h-mtTFA and its Saccharomyces cerevisiae homolog, sc-mtTFA. Wild-type sc-mtTFA is unable to activate transcription from the human mitochondrial light-strand promoter (LSP). Addition of the human tail region to sc-mtTFA conferred LSP-specific promoter activation. In all of the different h-mtTFA mutations tested, transcriptional activation was correlated with specific DNA-binding activity, suggesting that these two functions may be inseparable, a situation entirely consistent with previous mutational analyses of human mitochondrial promoters.
Many diverse taxa have evolved independently the habit of living in plant galls. For all but some viral galls, it is unknown whether plants produce galls as a specialized plant reaction to certain types of herbivory, or whether herbivores direct gall development. Here I present a phylogenetic analysis of gallforming cerataphidine aphids which demonstrates that gall morphology is extremely conservative with respect to aphid phylogeny, but variable with respect to plant taxonomy. In addition, the phylogeny reveals at least three host plant switches where the aphids produce galls most similar to the galls of their closest relatives, rather than galls similar to the galls of aphids already present on the host plant. These results suggest that aphids determine the details of gall morphology essentially extending their phenotype to include plant material. Based on this and other evidence, I suggest that the aphids and other galling insects manipulate latent plant developmental programmes to produce modified atavistic plant morphologies rather than create new forms de novo.