Filter
Associated Lab
- Aguilera Castrejon Lab (19) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (75) Apply Ahrens Lab filter
- Aso Lab (42) Apply Aso Lab filter
- Baker Lab (38) Apply Baker Lab filter
- Betzig Lab (116) Apply Betzig Lab filter
- Beyene Lab (15) Apply Beyene Lab filter
- Bock Lab (17) Apply Bock Lab filter
- Branson Lab (56) Apply Branson Lab filter
- Card Lab (43) Apply Card Lab filter
- Cardona Lab (64) Apply Cardona Lab filter
- Chklovskii Lab (13) Apply Chklovskii Lab filter
- Clapham Lab (16) Apply Clapham Lab filter
- Cui Lab (19) Apply Cui Lab filter
- Darshan Lab (12) Apply Darshan Lab filter
- Dennis Lab (3) Apply Dennis Lab filter
- Dickson Lab (46) Apply Dickson Lab filter
- Druckmann Lab (25) Apply Druckmann Lab filter
- Dudman Lab (58) Apply Dudman Lab filter
- Eddy/Rivas Lab (30) Apply Eddy/Rivas Lab filter
- Egnor Lab (11) Apply Egnor Lab filter
- Espinosa Medina Lab (25) Apply Espinosa Medina Lab filter
- Feliciano Lab (16) Apply Feliciano Lab filter
- Fetter Lab (41) Apply Fetter Lab filter
- FIB-SEM Technology (1) Apply FIB-SEM Technology filter
- Fitzgerald Lab (30) Apply Fitzgerald Lab filter
- Freeman Lab (15) Apply Freeman Lab filter
- Funke Lab (46) Apply Funke Lab filter
- Gonen Lab (91) Apply Gonen Lab filter
- Grigorieff Lab (62) Apply Grigorieff Lab filter
- Harris Lab (65) Apply Harris Lab filter
- Heberlein Lab (94) Apply Heberlein Lab filter
- Hermundstad Lab (32) Apply Hermundstad Lab filter
- Hess Lab (80) Apply Hess Lab filter
- Ilanges Lab (4) Apply Ilanges Lab filter
- Jayaraman Lab (49) Apply Jayaraman Lab filter
- Ji Lab (33) Apply Ji Lab filter
- Johnson Lab (7) Apply Johnson Lab filter
- Kainmueller Lab (19) Apply Kainmueller Lab filter
- Karpova Lab (15) Apply Karpova Lab filter
- Keleman Lab (13) Apply Keleman Lab filter
- Keller Lab (77) Apply Keller Lab filter
- Koay Lab (20) Apply Koay Lab filter
- Lavis Lab (162) Apply Lavis Lab filter
- Lee (Albert) Lab (34) Apply Lee (Albert) Lab filter
- Leonardo Lab (23) Apply Leonardo Lab filter
- Li Lab (32) Apply Li Lab filter
- Lippincott-Schwartz Lab (182) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (9) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (65) Apply Liu (Zhe) Lab filter
- Looger Lab (138) Apply Looger Lab filter
- Magee Lab (49) Apply Magee Lab filter
- Menon Lab (18) Apply Menon Lab filter
- Murphy Lab (13) Apply Murphy Lab filter
- O'Shea Lab (8) Apply O'Shea Lab filter
- Otopalik Lab (13) Apply Otopalik Lab filter
- Pachitariu Lab (56) Apply Pachitariu Lab filter
- Pastalkova Lab (19) Apply Pastalkova Lab filter
- Pavlopoulos Lab (19) Apply Pavlopoulos Lab filter
- Pedram Lab (15) Apply Pedram Lab filter
- Podgorski Lab (16) Apply Podgorski Lab filter
- Reiser Lab (55) Apply Reiser Lab filter
- Riddiford Lab (44) Apply Riddiford Lab filter
- Romani Lab (52) Apply Romani Lab filter
- Rubin Lab (149) Apply Rubin Lab filter
- Saalfeld Lab (66) Apply Saalfeld Lab filter
- Satou Lab (18) Apply Satou Lab filter
- Scheffer Lab (38) Apply Scheffer Lab filter
- Schreiter Lab (72) Apply Schreiter Lab filter
- Schulze Lab (1) Apply Schulze Lab filter
- Sgro Lab (23) Apply Sgro Lab filter
- Shroff Lab (35) Apply Shroff Lab filter
- Simpson Lab (23) Apply Simpson Lab filter
- Singer Lab (80) Apply Singer Lab filter
- Spruston Lab (98) Apply Spruston Lab filter
- Stern Lab (160) Apply Stern Lab filter
- Sternson Lab (54) Apply Sternson Lab filter
- Stringer Lab (44) Apply Stringer Lab filter
- Svoboda Lab (136) Apply Svoboda Lab filter
- Tebo Lab (36) Apply Tebo Lab filter
- Tervo Lab (10) Apply Tervo Lab filter
- Tillberg Lab (22) Apply Tillberg Lab filter
- Tjian Lab (64) Apply Tjian Lab filter
- Truman Lab (88) Apply Truman Lab filter
- Turaga Lab (53) Apply Turaga Lab filter
- Turner Lab (38) Apply Turner Lab filter
- Vale Lab (8) Apply Vale Lab filter
- Voigts Lab (5) Apply Voigts Lab filter
- Wang (Meng) Lab (31) Apply Wang (Meng) Lab filter
- Wang (Shaohe) Lab (25) Apply Wang (Shaohe) Lab filter
- Wong-Campos Lab (1) Apply Wong-Campos Lab filter
- Wu Lab (9) Apply Wu Lab filter
- Zlatic Lab (28) Apply Zlatic Lab filter
- Zuker Lab (25) Apply Zuker Lab filter
Associated Project Team
- CellMap (13) Apply CellMap filter
- COSEM (3) Apply COSEM filter
- FIB-SEM Technology (5) Apply FIB-SEM Technology filter
- Fly Descending Interneuron (12) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (14) Apply Fly Functional Connectome filter
- Fly Olympiad (5) Apply Fly Olympiad filter
- FlyEM (56) Apply FlyEM filter
- FlyLight (50) Apply FlyLight filter
- GENIE (47) Apply GENIE filter
- Integrative Imaging (11) Apply Integrative Imaging filter
- Larval Olympiad (2) Apply Larval Olympiad filter
- MouseLight (18) Apply MouseLight filter
- NeuroSeq (1) Apply NeuroSeq filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (29) Apply Tool Translation Team (T3) filter
- Transcription Imaging (49) Apply Transcription Imaging filter
Publication Date
- 2026 (118) Apply 2026 filter
- 2025 (223) Apply 2025 filter
- 2024 (209) Apply 2024 filter
- 2023 (158) Apply 2023 filter
- 2022 (192) Apply 2022 filter
- 2021 (194) Apply 2021 filter
- 2020 (196) Apply 2020 filter
- 2019 (202) Apply 2019 filter
- 2018 (232) Apply 2018 filter
- 2017 (217) Apply 2017 filter
- 2016 (209) Apply 2016 filter
- 2015 (252) Apply 2015 filter
- 2014 (236) Apply 2014 filter
- 2013 (194) Apply 2013 filter
- 2012 (190) Apply 2012 filter
- 2011 (190) Apply 2011 filter
- 2010 (161) Apply 2010 filter
- 2009 (158) Apply 2009 filter
- 2008 (140) Apply 2008 filter
- 2007 (106) Apply 2007 filter
- 2006 (92) Apply 2006 filter
- 2005 (67) Apply 2005 filter
- 2004 (57) Apply 2004 filter
- 2003 (58) Apply 2003 filter
- 2002 (39) Apply 2002 filter
- 2001 (28) Apply 2001 filter
- 2000 (29) Apply 2000 filter
- 1999 (14) Apply 1999 filter
- 1998 (18) Apply 1998 filter
- 1997 (16) Apply 1997 filter
- 1996 (10) Apply 1996 filter
- 1995 (18) Apply 1995 filter
- 1994 (12) Apply 1994 filter
- 1993 (10) Apply 1993 filter
- 1992 (6) Apply 1992 filter
- 1991 (11) Apply 1991 filter
- 1990 (11) Apply 1990 filter
- 1989 (6) Apply 1989 filter
- 1988 (1) Apply 1988 filter
- 1987 (7) Apply 1987 filter
- 1986 (4) Apply 1986 filter
- 1985 (5) Apply 1985 filter
- 1984 (2) Apply 1984 filter
- 1983 (2) Apply 1983 filter
- 1982 (3) Apply 1982 filter
- 1981 (3) Apply 1981 filter
- 1980 (1) Apply 1980 filter
- 1979 (1) Apply 1979 filter
- 1976 (2) Apply 1976 filter
- 1973 (1) Apply 1973 filter
- 1970 (1) Apply 1970 filter
- 1967 (1) Apply 1967 filter
Type of Publication
4313 Publications
Showing 2651-2660 of 4313 resultsMany motor control systems generate multiple movements using a common set of muscles. How are premotor circuits able to flexibly generate diverse movement patterns? Here, we characterize the neuronal circuits that drive the distinct courtship songs of Drosophila melanogaster. Male flies vibrate their wings towards females to produce two different song modes – pulse and sine song – which signal species identity and male quality. Using cell-type specific genetic reagents and the connectome, we provide a cellular and synaptic map of the circuits in the male ventral nerve cord that generate these songs and examine how activating or inhibiting each cell type within these circuits affects the song. Our data reveal that the song circuit is organized into two nested feed-forward pathways, with extensive reciprocal and feed-back connections. The larger network produces pulse song, the more complex and ancestral song form. A subset of this network produces sine song, the simpler and more recent form. Such nested organization may be a common feature of motor control circuits in which evolution has layered increasing flexibility on to a basic movement pattern.
In the cerebral cortex, local circuits consist of tens of thousands of neurons, each of which makes thousands of synaptic connections. Perhaps the biggest impediment to understanding these networks is that we have no wiring diagrams of their interconnections. Even if we had a partial or complete wiring diagram, however, understanding the network would also require information about each neuron’s function. Here we show that the relationship between structure and function can be studied in the cortex with a combination of in vivo physiology and network anatomy. We used two-photon calcium imaging to characterize a functional property–the preferred stimulus orientation–of a group of neurons in the mouse primary visual cortex. Large-scale electron microscopy of serial thin sections was then used to trace a portion of these neurons’ local network. Consistent with a prediction from recent physiological experiments, inhibitory interneurons received convergent anatomical input from nearby excitatory neurons with a broad range of preferred orientations, although weak biases could not be rejected.
Although hippocampal theta oscillations represent a prime example of temporal coding in the mammalian brain, little is known about the specific biophysical mechanisms. Intracellular recordings support a particular abstract oscillatory interference model of hippocampal theta activity, the soma-dendrite interference model. To gain insight into the cellular and circuit level mechanisms of theta activity, we implemented a similar form of interference using the actual hippocampal network in mice in vitro. We found that pairing increasing levels of phasic dendritic excitation with phasic stimulation of perisomatic projecting inhibitory interneurons induced a somatic polarization and action potential timing profile that reproduced most common features. Alterations in the temporal profile of inhibition were required to fully capture all features. These data suggest that theta-related place cell activity is generated through an interaction between a phasic dendritic excitation and a phasic perisomatic shunting inhibition delivered by interneurons, a subset of which undergo activity-dependent presynaptic modulation.
Complex networks are studied across many fields of science. To uncover their structural design principles, we defined “network motifs,” patterns of interconnections occurring in complex networks at numbers that are significantly higher than those in randomized networks. We found such motifs in networks from biochemistry, neurobiology, ecology, and engineering. The motifs shared by ecological food webs were distinct from the motifs shared by the genetic networks of Escherichia coli and Saccharomyces cerevisiae or from those found in the World Wide Web. Similar motifs were found in networks that perform information processing, even though they describe elements as different as biomolecules within a cell and synaptic connections between neurons in Caenorhabditis elegans. Motifs may thus define universal classes of networks. This approach may uncover the basic building blocks of most networks.
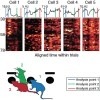
Regions within the prefrontal cortex are thought to process beliefs about the world, but little is known about the circuit dynamics underlying the formation and modification of these beliefs. Using a task that permits dissociation between the activity encoding an animal’s internal state and that encoding aspects of behavior, we found that transient increases in the volatility of activity in the rat medial prefrontal cortex accompany periods when an animal’s belief is modified after an environmental change. Activity across the majority of sampled neurons underwent marked, abrupt, and coordinated changes when prior belief was abandoned in favor of exploration of alternative strategies. These dynamics reflect network switches to a state of instability, which diminishes over the period of exploration as new stable representations are formed.
Brains comprise complex networks of neurons and connections, similar to the nodes and edges of artificial networks. Network analysis applied to the wiring diagrams of brains can offer insights into how they support computations and regulate the flow of information underlying perception and behaviour. The completion of the first whole-brain connectome of an adult fly, containing over 130,000 neurons and millions of synaptic connections, offers an opportunity to analyse the statistical properties and topological features of a complete brain. Here we computed the prevalence of two- and three-node motifs, examined their strengths, related this information to both neurotransmitter composition and cell type annotations, and compared these metrics with wiring diagrams of other animals. We found that the network of the fly brain displays rich-club organization, with a large population (30% of the connectome) of highly connected neurons. We identified subsets of rich-club neurons that may serve as integrators or broadcasters of signals. Finally, we examined subnetworks based on 78 anatomically defined brain regions or neuropils. These data products are shared within the FlyWire Codex (https://codex.flywire.ai) and should serve as a foundation for models and experiments exploring the relationship between neural activity and anatomical structure.
Visual, auditory, somatosensory, and olfactory stimuli generate temporally precise patterns of action potentials (spikes). It is unclear, however, how the precision of spike generation relates to the pattern and variability of synaptic input elicited by physiological stimuli. We determined how synaptic conductances evoked by light stimuli that activate the rod bipolar pathway control spike generation in three identified types of mouse retinal ganglion cells (RGCs). The relative amplitude, timing, and impact of excitatory and inhibitory input differed dramatically between On and Off RGCs. Spikes evoked by repeated somatic injection of identical light-evoked synaptic conductances were more temporally precise than those evoked by light. However, the precision of spikes evoked by conductances that varied from trial to trial was similar to that of light-evoked spikes. Thus, the rod bipolar pathway modulates different RGCs via unique combinations of synaptic input, and RGC temporal variability reflects variability in the input this circuit provides.
We give a covering number bound for deep learning networks that is independent of the size of the network. The key for the simple analysis is that for linear classifiers, rotating the data doesn't affect the covering number. Thus, we can ignore the rotation part of each layer's linear transformation, and get the covering number bound by concentrating on the scaling part.
Neural mechanisms underlying sexually dimorphic social behaviors remain enigmatic in most species. In Drosophila, sexually dimorphic P1/pC1x neurons have been described as a site of sensory integration that regulates mating and aggressive behaviors. We show that the male P1/pC1x population forms a highly intertwined network with male-specific mAL and aSP-a neurons that is poised to regulate male behavior. The 48 P1/pC1x cell types exhibit heterogeneous synaptic connections with a subset receiving strong input from identified sensory pathways. We also describe circuit motifs by which P1 and sexually dimorphic aIPg neurons co-regulate social behaviors. Genetic driver lines for these cell types were generated and used to discover distinct roles for P1/pC1x cell types in promoting social acoustic signaling and male-male interactions. Our results reveal unexpected diversity in the connectivity and behavioral roles of the P1/pC1x cell types and provide essential genetic tools for interrogating their neurophysiological and behavioral functions.
Due to advances in electron microscopy and deep learning, it is now practical to reconstruct a connectome, a description of neurons and the chemical synapses between them, for significant volumes of neural tissue. Smaller past reconstructions were primarily used by domain experts, could be handled by downloading data, and performance was not a serious problem. But new and much larger reconstructions upend these assumptions. These networks now contain tens of thousands of neurons and tens of millions of connections, with yet larger reconstructions pending, and are of interest to a large community of non-specialists. Allowing other scientists to make use of this data needs more than publication-it requires new tools that are publicly available, easy to use, and efficiently handle large data. We introduce neuPrint to address these data analysis challenges. Neuprint contains two major components-a web interface and programmer APIs. The web interface is designed to allow any scientist worldwide, using only a browser, to quickly ask and answer typical biological queries about a connectome. The neuPrint APIs allow more computer-savvy scientists to make more complex or higher volume queries. NeuPrint also provides features for assessing reconstruction quality. Internally, neuPrint organizes connectome data as a graph stored in a neo4j database. This gives high performance for typical queries, provides access though a public and well documented query language Cypher, and will extend well to future larger connectomics databases. Our experience is also an experiment in open science. We find a significant fraction of the readers of the article proceed to examine the data directly. In our case preprints worked exactly as intended, with data inquiries and PDF downloads starting immediately after pre-print publication, and little affected by formal publication later. From this we deduce that many readers are more interested in our data than in our analysis of our data, suggesting that data-only papers can be well appreciated and that public data release can speed up the propagation of scientific results by many months. We also find that providing, and keeping, the data available for online access imposes substantial additional costs to connectomics research.