Filter
Associated Lab
- Branson Lab (4) Apply Branson Lab filter
- Card Lab (5) Apply Card Lab filter
- Funke Lab (1) Apply Funke Lab filter
- Jayaraman Lab (2) Apply Jayaraman Lab filter
- Remove Reiser Lab filter Reiser Lab
- Romani Lab (4) Apply Romani Lab filter
- Rubin Lab (15) Apply Rubin Lab filter
- Stern Lab (1) Apply Stern Lab filter
- Turaga Lab (3) Apply Turaga Lab filter
- Zuker Lab (1) Apply Zuker Lab filter
Associated Project Team
Publication Date
- 2025 (3) Apply 2025 filter
- 2024 (2) Apply 2024 filter
- 2023 (6) Apply 2023 filter
- 2022 (3) Apply 2022 filter
- 2021 (3) Apply 2021 filter
- 2020 (2) Apply 2020 filter
- 2019 (2) Apply 2019 filter
- 2018 (4) Apply 2018 filter
- 2017 (6) Apply 2017 filter
- 2016 (3) Apply 2016 filter
- 2015 (2) Apply 2015 filter
- 2014 (2) Apply 2014 filter
- 2013 (1) Apply 2013 filter
- 2012 (1) Apply 2012 filter
- 2011 (2) Apply 2011 filter
- 2010 (2) Apply 2010 filter
- 2009 (1) Apply 2009 filter
Type of Publication
- Remove Janelia filter Janelia
45 Publications
Showing 31-40 of 45 resultsThe delivery of tracers into populations of neurons is essential to visualize their anatomy and analyze their function. In some model systems genetically-targeted expression of fluorescent proteins is the method of choice; however, these genetic tools are not available for most organisms and alternative labeling methods are very limited. Here we describe a new method for neuronal labelling by electrophoretic dye delivery from a suction electrode directly through the neuronal sheath of nerves and ganglia in insects. Polar tracer molecules were delivered into the locust auditory nerve without destroying its function, simultaneously staining peripheral sensory structures and central axonal projections. Local neuron populations could be labelled directly through the surface of the brain, and in-vivo optical imaging of sound-evoked activity was achieved through the electrophoretic delivery of calcium indicators. The method provides a new tool for studying how stimuli are processed in peripheral and central sensory pathways and is a significant advance for the study of nervous systems in non-model organisms.
Visual projection neurons (VPNs) provide an anatomical connection between early visual processing and higher brain regions. Here we characterize lobula columnar (LC) cells, a class of Drosophila VPNs that project to distinct central brain structures called optic glomeruli. We anatomically describe 22 different LC types and show that, for several types, optogenetic activation in freely moving flies evokes specific behaviors. The activation phenotypes of two LC types closely resemble natural avoidance behaviors triggered by a visual loom. In vivo two-photon calcium imaging reveals that these LC types respond to looming stimuli, while another type does not, but instead responds to the motion of a small object. Activation of LC neurons on only one side of the brain can result in attractive or aversive turning behaviors depending on the cell type. Our results indicate that LC neurons convey information on the presence and location of visual features relevant for specific behaviors.
Color is famous for not existing in the external world: our brains create the perception of color from the spatial and temporal patterns of the wavelength and intensity of light. For an intangible quality, we have detailed knowledge of its origins and consequences. Much is known about the organization and evolution of the first phases of color processing, the filtering of light in the eye and processing in the retina, and about the final phases, the roles of color in behavior and natural selection. To understand how color processing in the central brain has evolved, we need well-defined pathways or circuitry where we can gauge how color contributes to the computations involved in specific behaviors. Examples of such pathways or circuitry that are dedicated to processing color cues are rare, despite the separation of color and luminance pathways early in the visual system of many species, and despite the traditional definition of color as being independent of luminance. This minireview presents examples in which color vision contributes to behaviors dominated by other visual modalities, examples that are not part of the canon of color vision circuitry. The pathways and circuitry process a range of chromatic properties of objects and their illumination, and are taken from a variety of species. By considering how color processing complements luminance processing, rather than being independent of it, we gain an additional way to account for the diversity of color coding in the central brain, its consequences for specific behaviors and ultimately the evolution of color vision.
Animal species display enormous variation for innate behaviours, but little is known about how this diversity arose. Here, using an unbiased genetic approach, we map a courtship song difference between wild isolates of Drosophila simulans and Drosophila mauritiana to a 966 base pair region within the slowpoke (slo) locus, which encodes a calcium-activated potassium channel. Using the reciprocal hemizygosity test, we confirm that slo is the causal locus and resolve the causal mutation to the evolutionarily recent insertion of a retroelement in a slo intron within D. simulans. Targeted deletion of this retroelement reverts the song phenotype and alters slo splicing. Like many ion channel genes, slo is expressed widely in the nervous system and influences a variety of behaviours; slo-null males sing little song with severely disrupted features. By contrast, the natural variant of slo alters a specific component of courtship song, illustrating that regulatory evolution of a highly pleiotropic ion channel gene can cause modular changes in behaviour.
Animal sensory systems are optimally adapted to those features typically encountered in natural surrounds, thus allowing neurons with limited bandwidth to encode challengingly large input ranges. Natural scenes are not random, and peripheral visual systems in vertebrates and insects have evolved to respond efficiently to their typical spatial statistics. The mammalian visual cortex is also tuned to natural spatial statistics, but less is known about coding in higher order neurons in insects. To redress this we here record intracellularly from a higher order visual neuron in the hoverfly. We show that the cSIFE neuron, which is inhibited by stationary images, is maximally inhibited when the slope constant of the amplitude spectrum is close to the mean in natural scenes. The behavioural optomotor response is also strongest to images with naturalistic image statistics. Our results thus reveal a close coupling between the inherent statistics of natural scenes and higher order visual processing in insects.

An important strategy for efficient neural coding is to match the range of cellular responses to the distribution of relevant input signals. However, the structure and relevance of sensory signals depend on behavioral state. Here, we show that behavior modifies neural activity at the earliest stages of fly vision. We describe a class of wide-field neurons that provide feedback to the most peripheral layer of the Drosophila visual system, the lamina. Using in vivo patch-clamp electrophysiology, we found that lamina wide-field neurons respond to low-frequency luminance fluctuations. Recordings in flying flies revealed that the gain and frequency tuning of wide-field neurons change during flight, and that these effects are mimicked by the neuromodulator octopamine. Genetically silencing wide-field neurons increased behavioral responses to slow-motion stimuli. Together, these findings identify a cell type that is gated by behavior to enhance neural coding by subtracting low-frequency signals from the inputs to motion detection circuits.

Visual motion perception is critical to many animal behaviors, and flies have emerged as a powerful model system for exploring this fundamental neural computation. Although numerous studies have suggested that fly motion vision is governed by a simple neural circuit [1-3], the implementation of this circuit has remained mysterious for decades. Connectomics and neurogenetics have produced a surge in recent progress, and several studies have shown selectivity for light increments (ON) or decrements (OFF) in key elements associated with this circuit [4-7]. However, related studies have reached disparate conclusions about where this selectivity emerges and whether it plays a major role in motion vision [8-13]. To address these questions, we examined activity in the neuropil thought to be responsible for visual motion detection, the medulla, of Drosophila melanogaster in response to a range of visual stimuli using two-photon calcium imaging. We confirmed that the input neurons of the medulla, the LMCs, are not responsible for light-on and light-off selectivity. We then examined the pan-neural response of medulla neurons and found prominent selectivity for light-on and light-off in layers of the medulla associated with two anatomically derived pathways (L1/L2 associated) [14, 15]. We next examined the activity of prominent interneurons within each pathway (Mi1 and Tm1) and found that these neurons have corresponding selectivity for light-on or light-off. These results provide direct evidence that motion is computed in parallel light-on and light-off pathways, demonstrate that this selectivity emerges in neurons immediately downstream of the LMCs, and specify where crucial elements of motion computation occur.
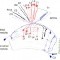
Motion detection is a fundamental neural computation performed by many sensory systems. In the fly, local motion computation is thought to occur within the first two layers of the visual system, the lamina and medulla. We constructed specific genetic driver lines for each of the 12 neuron classes in the lamina. We then depolarized and hyperpolarized each neuron type and quantified fly behavioral responses to a diverse set of motion stimuli. We found that only a small number of lamina output neurons are essential for motion detection, while most neurons serve to sculpt and enhance these feedforward pathways. Two classes of feedback neurons (C2 and C3), and lamina output neurons (L2 and L4), are required for normal detection of directional motion stimuli. Our results reveal a prominent role for feedback and lateral interactions in motion processing and demonstrate that motion-dependent behaviors rely on contributions from nearly all lamina neuron classes.

Virtual reality (VR) holds great promise as a tool to study the neural circuitry underlying animal behaviors. Here, we discuss the advantages of VR and the experimental paradigms and technologies that enable closed loop behavioral experiments. We review recent results from VR research in genetic model organisms where the potential combination of rich behaviors, genetic tools and cutting edge neural recording techniques are leading to breakthroughs in our understanding of the neural basis of behavior. We also discuss several key issues to consider when performing VR experiments and provide an outlook for the future of this exciting experimental toolkit.