Filter
Associated Lab
- Aguilera Castrejon Lab (16) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (63) Apply Ahrens Lab filter
- Aso Lab (40) Apply Aso Lab filter
- Baker Lab (38) Apply Baker Lab filter
- Betzig Lab (112) Apply Betzig Lab filter
- Beyene Lab (13) Apply Beyene Lab filter
- Bock Lab (17) Apply Bock Lab filter
- Branson Lab (52) Apply Branson Lab filter
- Card Lab (41) Apply Card Lab filter
- Cardona Lab (63) Apply Cardona Lab filter
- Chklovskii Lab (13) Apply Chklovskii Lab filter
- Clapham Lab (14) Apply Clapham Lab filter
- Cui Lab (19) Apply Cui Lab filter
- Darshan Lab (12) Apply Darshan Lab filter
- Dennis Lab (1) Apply Dennis Lab filter
- Dickson Lab (46) Apply Dickson Lab filter
- Druckmann Lab (25) Apply Druckmann Lab filter
- Dudman Lab (50) Apply Dudman Lab filter
- Eddy/Rivas Lab (30) Apply Eddy/Rivas Lab filter
- Egnor Lab (11) Apply Egnor Lab filter
- Espinosa Medina Lab (19) Apply Espinosa Medina Lab filter
- Feliciano Lab (7) Apply Feliciano Lab filter
- Fetter Lab (41) Apply Fetter Lab filter
- Fitzgerald Lab (29) Apply Fitzgerald Lab filter
- Freeman Lab (15) Apply Freeman Lab filter
- Funke Lab (38) Apply Funke Lab filter
- Gonen Lab (91) Apply Gonen Lab filter
- Grigorieff Lab (62) Apply Grigorieff Lab filter
- Harris Lab (60) Apply Harris Lab filter
- Heberlein Lab (94) Apply Heberlein Lab filter
- Hermundstad Lab (26) Apply Hermundstad Lab filter
- Hess Lab (77) Apply Hess Lab filter
- Ilanges Lab (2) Apply Ilanges Lab filter
- Jayaraman Lab (46) Apply Jayaraman Lab filter
- Ji Lab (33) Apply Ji Lab filter
- Johnson Lab (6) Apply Johnson Lab filter
- Kainmueller Lab (19) Apply Kainmueller Lab filter
- Karpova Lab (14) Apply Karpova Lab filter
- Keleman Lab (13) Apply Keleman Lab filter
- Keller Lab (76) Apply Keller Lab filter
- Koay Lab (18) Apply Koay Lab filter
- Lavis Lab (148) Apply Lavis Lab filter
- Lee (Albert) Lab (34) Apply Lee (Albert) Lab filter
- Leonardo Lab (23) Apply Leonardo Lab filter
- Li Lab (28) Apply Li Lab filter
- Lippincott-Schwartz Lab (168) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (6) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (61) Apply Liu (Zhe) Lab filter
- Looger Lab (138) Apply Looger Lab filter
- Magee Lab (49) Apply Magee Lab filter
- Menon Lab (18) Apply Menon Lab filter
- Murphy Lab (13) Apply Murphy Lab filter
- O'Shea Lab (6) Apply O'Shea Lab filter
- Otopalik Lab (13) Apply Otopalik Lab filter
- Pachitariu Lab (47) Apply Pachitariu Lab filter
- Pastalkova Lab (18) Apply Pastalkova Lab filter
- Pavlopoulos Lab (19) Apply Pavlopoulos Lab filter
- Pedram Lab (15) Apply Pedram Lab filter
- Podgorski Lab (16) Apply Podgorski Lab filter
- Reiser Lab (51) Apply Reiser Lab filter
- Riddiford Lab (44) Apply Riddiford Lab filter
- Romani Lab (43) Apply Romani Lab filter
- Rubin Lab (143) Apply Rubin Lab filter
- Saalfeld Lab (63) Apply Saalfeld Lab filter
- Satou Lab (16) Apply Satou Lab filter
- Scheffer Lab (36) Apply Scheffer Lab filter
- Schreiter Lab (67) Apply Schreiter Lab filter
- Sgro Lab (21) Apply Sgro Lab filter
- Shroff Lab (30) Apply Shroff Lab filter
- Simpson Lab (23) Apply Simpson Lab filter
- Singer Lab (80) Apply Singer Lab filter
- Spruston Lab (93) Apply Spruston Lab filter
- Stern Lab (156) Apply Stern Lab filter
- Sternson Lab (54) Apply Sternson Lab filter
- Stringer Lab (35) Apply Stringer Lab filter
- Svoboda Lab (135) Apply Svoboda Lab filter
- Tebo Lab (33) Apply Tebo Lab filter
- Tervo Lab (9) Apply Tervo Lab filter
- Tillberg Lab (21) Apply Tillberg Lab filter
- Tjian Lab (64) Apply Tjian Lab filter
- Truman Lab (88) Apply Truman Lab filter
- Turaga Lab (50) Apply Turaga Lab filter
- Turner Lab (37) Apply Turner Lab filter
- Vale Lab (7) Apply Vale Lab filter
- Voigts Lab (3) Apply Voigts Lab filter
- Wang (Meng) Lab (18) Apply Wang (Meng) Lab filter
- Wang (Shaohe) Lab (25) Apply Wang (Shaohe) Lab filter
- Wu Lab (9) Apply Wu Lab filter
- Zlatic Lab (28) Apply Zlatic Lab filter
- Zuker Lab (25) Apply Zuker Lab filter
Associated Project Team
- CellMap (12) Apply CellMap filter
- COSEM (3) Apply COSEM filter
- FIB-SEM Technology (2) Apply FIB-SEM Technology filter
- Fly Descending Interneuron (10) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (14) Apply Fly Functional Connectome filter
- Fly Olympiad (5) Apply Fly Olympiad filter
- FlyEM (53) Apply FlyEM filter
- FlyLight (49) Apply FlyLight filter
- GENIE (45) Apply GENIE filter
- Integrative Imaging (3) Apply Integrative Imaging filter
- Larval Olympiad (2) Apply Larval Olympiad filter
- MouseLight (18) Apply MouseLight filter
- NeuroSeq (1) Apply NeuroSeq filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (26) Apply Tool Translation Team (T3) filter
- Transcription Imaging (49) Apply Transcription Imaging filter
Publication Date
- 2025 (92) Apply 2025 filter
- 2024 (221) Apply 2024 filter
- 2023 (160) Apply 2023 filter
- 2022 (193) Apply 2022 filter
- 2021 (194) Apply 2021 filter
- 2020 (196) Apply 2020 filter
- 2019 (202) Apply 2019 filter
- 2018 (232) Apply 2018 filter
- 2017 (217) Apply 2017 filter
- 2016 (209) Apply 2016 filter
- 2015 (252) Apply 2015 filter
- 2014 (236) Apply 2014 filter
- 2013 (194) Apply 2013 filter
- 2012 (190) Apply 2012 filter
- 2011 (190) Apply 2011 filter
- 2010 (161) Apply 2010 filter
- 2009 (158) Apply 2009 filter
- 2008 (140) Apply 2008 filter
- 2007 (106) Apply 2007 filter
- 2006 (92) Apply 2006 filter
- 2005 (67) Apply 2005 filter
- 2004 (57) Apply 2004 filter
- 2003 (58) Apply 2003 filter
- 2002 (39) Apply 2002 filter
- 2001 (28) Apply 2001 filter
- 2000 (29) Apply 2000 filter
- 1999 (14) Apply 1999 filter
- 1998 (18) Apply 1998 filter
- 1997 (16) Apply 1997 filter
- 1996 (10) Apply 1996 filter
- 1995 (18) Apply 1995 filter
- 1994 (12) Apply 1994 filter
- 1993 (10) Apply 1993 filter
- 1992 (6) Apply 1992 filter
- 1991 (11) Apply 1991 filter
- 1990 (11) Apply 1990 filter
- 1989 (6) Apply 1989 filter
- 1988 (1) Apply 1988 filter
- 1987 (7) Apply 1987 filter
- 1986 (4) Apply 1986 filter
- 1985 (5) Apply 1985 filter
- 1984 (2) Apply 1984 filter
- 1983 (2) Apply 1983 filter
- 1982 (3) Apply 1982 filter
- 1981 (3) Apply 1981 filter
- 1980 (1) Apply 1980 filter
- 1979 (1) Apply 1979 filter
- 1976 (2) Apply 1976 filter
- 1973 (1) Apply 1973 filter
- 1970 (1) Apply 1970 filter
- 1967 (1) Apply 1967 filter
Type of Publication
4079 Publications
Showing 1781-1790 of 4079 resultsHox genes encode highly conserved transcription factors that regionalize the animal body axis by controlling complex developmental processes. Although they are known to operate in multiple cell types and at different stages, we are still missing the batteries of genes targeted by any one Hox gene over the course of a single developmental process to achieve a particular cell and organ morphology. The transformation of wings into halteres by the Hox gene Ultrabithorax (Ubx) in Drosophila melanogaster presents an excellent model system to study the Hox control of transcriptional networks during successive stages of appendage morphogenesis and cell differentiation. We have used an inducible misexpression system to switch on Ubx in the wing epithelium at successive stages during metamorphosis–in the larva, prepupa, and pupa. We have then used extensive microarray expression profiling and quantitative RT-PCR to identify the primary transcriptional responses to Ubx. We find that Ubx targets range from regulatory genes like transcription factors and signaling components to terminal differentiation genes affecting a broad repertoire of cell behaviors and metabolic reactions. Ubx up- and down-regulates hundreds of downstream genes at each stage, mostly in a subtle manner. Strikingly, our analysis reveals that Ubx target genes are largely distinct at different stages of appendage morphogenesis, suggesting extensive interactions between Hox genes and hormone-controlled regulatory networks to orchestrate complex genetic programs during metamorphosis.
Genetic studies of the targets of the Hox genes have revealed only the tip of the iceberg. Recent microarray studies that have identified hundreds more transcriptional responses to Hox genes in Drosophila will help elucidate the role of Hox genes in development and evolution.
Neural development requires both synapse elaboration and elimination, yet relatively little is known about how these opposing activities are coordinated. Here, we provide evidence Hts/Adducin can serve this function. We show that Drosophila Hts/Adducin is enriched both pre- and postsynaptically at the NMJ. We then demonstrate that presynaptic Hts/Adducin is necessary and sufficient to control two opposing processes associated with synapse remodeling: (1) synapse stabilization as determined by light level and ultrastructural and electrophysiological assays and (2) the elaboration of actin-based, filopodia-like protrusions that drive synaptogenesis and growth. Synapse remodeling is sensitive to Hts/Adducin levels, and we provide evidence that the synaptic localization of Hts/Adducin is controlled via phosphorylation. Mechanistically, Drosophila Hts/Adducin protein has actin-capping activity. We propose that phosphorylation-dependent regulation of Hts/Adducin controls the level, localization, and activity of Hts/Adducin, influencing actin-based synapse elaboration and spectrin-based synapse stabilization. Hts/Adducin may define a mechanism to switch between synapse stability and dynamics.
In the perception of color, wavelengths of light reflected off objects are transformed into the derived quantities of brightness, saturation and hue. Neurons responding selectively to hue have been reported in primate cortex, but it is unknown how their narrow tuning in color space is produced by upstream circuit mechanisms. We report the discovery of neurons in the Drosophila optic lobe with hue-selective properties, which enables circuit-level analysis of color processing. From our analysis of an electron microscopy volume of a whole Drosophila brain, we construct a connectomics-constrained circuit model that accounts for this hue selectivity. Our model predicts that recurrent connections in the circuit are critical for generating hue selectivity. Experiments using genetic manipulations to perturb recurrence in adult flies confirm this prediction. Our findings reveal a circuit basis for hue selectivity in color vision.
Activation of gene transcription in mammalian cells requires several classes of coactivators that participate in different steps of the activation cascade. Using conventional and affinity chromatography, we have isolated a human coactivator complex that interacts directly with the C-terminal domain (CTD) of RNA polymerase II (Pol II). The CTD-binding complex is structurally and functionally indistinguishable from our previously isolated CRSP coactivator complex. The closely related, but transcriptionally inactive, ARC-L complex failed to interact with the CTD, indicating a significant biochemical difference between CRSP and ARC-L that may, in part, explain their functional divergence. Electron microscopy and three-dimensional single-particle reconstruction reveals a conformation for CTD-CRSP that is structurally distinct from unliganded CRSP or CRSP bound to SREBP-1a, but highly similar to CRSP bound to the VP16 activator. Together, our findings suggest that the human CRSP coactivator functions, at least in part, by mediating activator-dependent recruitment of RNA Pol II via the CTD.
The recent derivation of human trophoblast stem cells (TSCs) from placental cytotrophoblasts and blastocysts opened opportunities for studying the development and function of the human placenta. Recent reports have suggested that human naïve, but not primed, pluripotent stem cells (PSCs) retain an exclusive potential to generate TSCs. Here we report that, in the absence of WNT stimulation, transforming growth factor β (TGF-β) pathway inhibition leads to direct and robust conversion of primed human PSCs into TSCs. The resulting primed PSC-derived TSC lines exhibit self-renewal, can differentiate into the main trophoblast lineages, and present RNA and epigenetic profiles that are indistinguishable from recently established TSC lines derived from human placenta, blastocysts, or isogenic human naïve PSCs expanded under human enhanced naïve stem cell medium (HENSM) conditions. Activation of nuclear Yes-associated protein (YAP) signaling is sufficient for this conversion and necessary for human TSC maintenance. Our findings underscore a residual plasticity in primed human PSCs that allows their in vitro conversion into extra-embryonic trophoblast lineages.
Physiological need states direct decision-making toward re-establishing homeostasis. Using a two-alternative forced choice task for mice that models elements of human decisions, we found that varying hunger and thirst states caused need-inappropriate choices, such as food seeking when thirsty. These results show limits on interoceptive knowledge of hunger and thirst states to guide decision-making. Instead, need states were identified after food and water consumption by outcome evaluation, which depended on the medial prefrontal cortex.
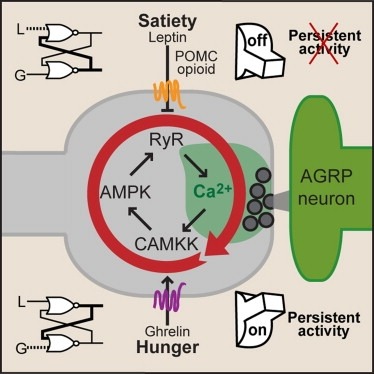
Synaptic plasticity in response to changes in physiologic state is coordinated by hormonal signals across multiple neuronal cell types. Here, we combine cell-type-specific electrophysiological, pharmacological, and optogenetic techniques to dissect neural circuits and molecular pathways controlling synaptic plasticity onto AGRP neurons, a population that regulates feeding. We find that food deprivation elevates excitatory synaptic input, which is mediated by a presynaptic positive feedback loop involving AMP-activated protein kinase. Potentiation of glutamate release was triggered by the orexigenic hormone ghrelin and exhibited hysteresis, persisting for hours after ghrelin removal. Persistent activity was reversed by the anorexigenic hormone leptin, and optogenetic photostimulation demonstrated involvement of opioid release from POMC neurons. Based on these experiments, we propose a memory storage device for physiological state constructed from bistable synapses that are flipped between two sustained activity states by transient exposure to hormones signaling energy levels.