Filter
Associated Lab
- Ahrens Lab (4) Apply Ahrens Lab filter
- Betzig Lab (2) Apply Betzig Lab filter
- Branson Lab (1) Apply Branson Lab filter
- Darshan Lab (3) Apply Darshan Lab filter
- Druckmann Lab (5) Apply Druckmann Lab filter
- Dudman Lab (3) Apply Dudman Lab filter
- Fetter Lab (1) Apply Fetter Lab filter
- Freeman Lab (3) Apply Freeman Lab filter
- Harris Lab (6) Apply Harris Lab filter
- Hermundstad Lab (1) Apply Hermundstad Lab filter
- Jayaraman Lab (9) Apply Jayaraman Lab filter
- Ji Lab (2) Apply Ji Lab filter
- Karpova Lab (1) Apply Karpova Lab filter
- Lavis Lab (4) Apply Lavis Lab filter
- Lee (Albert) Lab (3) Apply Lee (Albert) Lab filter
- Leonardo Lab (2) Apply Leonardo Lab filter
- Liu (Zhe) Lab (1) Apply Liu (Zhe) Lab filter
- Looger Lab (20) Apply Looger Lab filter
- Magee Lab (1) Apply Magee Lab filter
- Pachitariu Lab (2) Apply Pachitariu Lab filter
- Podgorski Lab (2) Apply Podgorski Lab filter
- Romani Lab (5) Apply Romani Lab filter
- Rubin Lab (3) Apply Rubin Lab filter
- Saalfeld Lab (2) Apply Saalfeld Lab filter
- Schreiter Lab (13) Apply Schreiter Lab filter
- Spruston Lab (3) Apply Spruston Lab filter
- Sternson Lab (4) Apply Sternson Lab filter
- Remove Svoboda Lab filter Svoboda Lab
- Tillberg Lab (3) Apply Tillberg Lab filter
- Turner Lab (3) Apply Turner Lab filter
Associated Project Team
Associated Support Team
- Anatomy and Histology (4) Apply Anatomy and Histology filter
- Gene Targeting and Transgenics (3) Apply Gene Targeting and Transgenics filter
- Integrative Imaging (2) Apply Integrative Imaging filter
- Janelia Experimental Technology (4) Apply Janelia Experimental Technology filter
- Molecular Genomics (2) Apply Molecular Genomics filter
- Primary & iPS Cell Culture (2) Apply Primary & iPS Cell Culture filter
- Project Technical Resources (1) Apply Project Technical Resources filter
- Quantitative Genomics (1) Apply Quantitative Genomics filter
- Scientific Computing Software (2) Apply Scientific Computing Software filter
- Viral Tools (2) Apply Viral Tools filter
Publication Date
- 2023 (5) Apply 2023 filter
- 2022 (6) Apply 2022 filter
- 2021 (7) Apply 2021 filter
- 2020 (5) Apply 2020 filter
- 2019 (14) Apply 2019 filter
- 2018 (11) Apply 2018 filter
- 2017 (9) Apply 2017 filter
- 2016 (8) Apply 2016 filter
- 2015 (9) Apply 2015 filter
- 2014 (7) Apply 2014 filter
- 2013 (10) Apply 2013 filter
- 2012 (9) Apply 2012 filter
- 2011 (7) Apply 2011 filter
- 2010 (7) Apply 2010 filter
- 2009 (8) Apply 2009 filter
- 2008 (6) Apply 2008 filter
- 2007 (3) Apply 2007 filter
131 Janelia Publications
Showing 51-60 of 131 resultsTremendous progress has been made since Neuron published our Primer on genetic dissection of neural circuits 10 years ago. Since then, cell-type-specific anatomical, neurophysiological, and perturbation studies have been carried out in a multitude of invertebrate and vertebrate organisms, linking neurons and circuits to behavioral functions. New methods allow systematic classification of cell types and provide genetic access to diverse neuronal types for studies of connectivity and neural coding during behavior. Here we evaluate key advances over the past decade and discuss future directions.
Neuroscience research is becoming increasingly more collaborative and interdisciplinary with partnerships between industry and academia and insights from fields beyond neuroscience. In the age of institutional initiatives and multi-investigator collaborations, scientists from around the world shared their perspectives on the effectiveness of large-scale collaborations versus single-lab, hypothesis-driven science.
Two-photon microscopy together with fluorescent proteins and fluorescent protein-based biosensors are commonly used tools in neuroscience. To enhance their experimental scope, it is important to optimize fluorescent proteins for two-photon excitation. Directed evolution of fluorescent proteins under one-photon excitation is common, but many one-photon properties do not correlate with two-photon properties. A simple system for expressing fluorescent protein mutants is colonies on an agar plate. The small focal volume of two-photon excitation makes creating a high throughput screen in this system a challenge for a conventional point-scanning approach. We present an instrument and accompanying software that solves this challenge by selectively scanning each colony based on a colony map captured under one-photon excitation. This instrument, called the GIZMO, can measure the two-photon excited fluorescence of 10,000 colonies in 7 hours. We show that the GIZMO can be used to evolve a fluorescent protein under two-photon excitation.
The ability to probe the membrane potential of multiple genetically defined neurons simultaneously would have a profound impact on neuroscience research. Genetically encoded voltage indicators are a promising tool for this purpose, and recent developments have achieved a high signal-to-noise ratio in vivo with 1-photon fluorescence imaging. However, these recordings exhibit several sources of noise and signal extraction remains a challenge. We present an improved signal extraction pipeline, spike-guided penalized matrix decomposition-nonnegative matrix factorization (SGPMD-NMF), which resolves supra- and subthreshold voltages in vivo. The method incorporates biophysical and optical constraints. We validate the pipeline with simultaneous patch-clamp and optical recordings from mouse layer 1 in vivo and with simulated and composite datasets with realistic noise. We demonstrate applications to mouse hippocampus expressing paQuasAr3-s or SomArchon1, mouse cortex expressing SomArchon1 or Voltron, and zebrafish spines expressing zArchon1.
Calcium imaging with genetically encoded calcium indicators (GECIs) is routinely used to measure neural activity in intact nervous systems. GECIs are frequently used in one of two different modes: to track activity in large populations of neuronal cell bodies, or to follow dynamics in subcellular compartments such as axons, dendrites and individual synaptic compartments. Despite major advances, calcium imaging is still limited by the biophysical properties of existing GECIs, including affinity, signal-to-noise ratio, rise and decay kinetics and dynamic range. Using structure-guided mutagenesis and neuron-based screening, we optimized the green fluorescent protein-based GECI GCaMP6 for different modes of in vivo imaging. The resulting jGCaMP7 sensors provide improved detection of individual spikes (jGCaMP7s,f), imaging in neurites and neuropil (jGCaMP7b), and may allow tracking larger populations of neurons using two-photon (jGCaMP7s,f) or wide-field (jGCaMP7c) imaging.
Calcium imaging with genetically encoded calcium indicators (GECIs) is routinely used to measure neural activity in intact nervous systems. GECIs are frequently used in one of two different modes: to track activity in large populations of neuronal cell bodies, or to follow dynamics in subcellular compartments such as axons, dendrites and individual synaptic compartments. Despite major advances, calcium imaging is still limited by the biophysical properties of existing GECIs, including affinity, signal-to-noise ratio, rise and decay kinetics, and dynamic range. Using structure-guided mutagenesis and neuron-based screening, we optimized the green fluorescent protein-based GECI GCaMP6 for different modes of in vivo imaging. The jGCaMP7 sensors provide improved detection of individual spikes (jGCaMP7s,f), imaging in neurites and neuropil (jGCaMP7b), and tracking large populations of neurons using 2-photon (jGCaMP7s,f) or wide-field (jGCaMP7c) imaging.
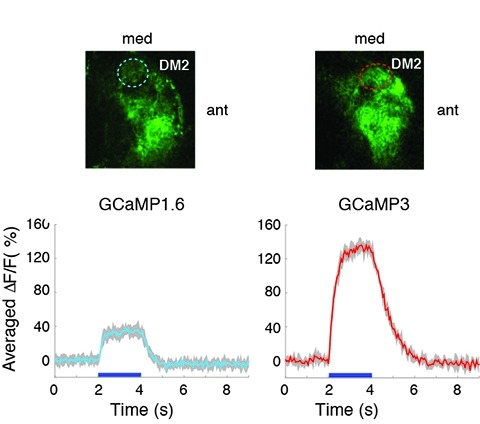
Genetically encoded calcium indicators (GECIs) can be used to image activity in defined neuronal populations. However, current GECIs produce inferior signals compared to synthetic indicators and recording electrodes, precluding detection of low firing rates. We developed a single-wavelength GCaMP2-based GECI (GCaMP3), with increased baseline fluorescence (3-fold), increased dynamic range (3-fold) and higher affinity for calcium (1.3-fold). We detected GCaMP3 fluorescence changes triggered by single action potentials in pyramidal cell dendrites, with signal-to-noise ratio and photostability substantially better than those of GCaMP2, D3cpVenus and TN-XXL. In Caenorhabditis elegans chemosensory neurons and the Drosophila melanogaster antennal lobe, sensory stimulation-evoked fluorescence responses were significantly enhanced with GCaMP3 (4-6-fold). In somatosensory and motor cortical neurons in the intact mouse, GCaMP3 detected calcium transients with amplitudes linearly dependent on action potential number. Long-term imaging in the motor cortex of behaving mice revealed large fluorescence changes in imaged neurons over months.
The basal ganglia play a critical role in the regulation of voluntary action in vertebrates. Our understanding of the function of the basal ganglia relies heavily upon anatomical information, but continued progress will require an understanding of the specific functional roles played by diverse cell types and their connectivity. An increasing number of mouse lines allow extensive identification, characterization, and manipulation of specified cell types in the basal ganglia. Despite the promise of genetically modified mice for elucidating the functional roles of diverse cell types, there is relatively little anatomical data obtained directly in the mouse. Here we have characterized the retrograde labeling obtained from a series of tracer injections throughout the dorsal striatum of adult mice. We found systematic variations in input along both the medial-lateral and anterior-posterior neuraxes in close agreement with canonical features of basal ganglia anatomy in the rat. In addition to the canonical features we have provided experimental support for the importance of non-canonical inputs to the striatum from the raphe nuclei and the amygdala. To look for organization at a finer scale we have analyzed the correlation structure of labeling intensity across our entire dataset. Using this analysis we found substantial local heterogeneity within the large-scale order. From this analysis we conclude that individual striatal sites receive varied combinations of cortical and thalamic input from multiple functional areas, consistent with some earlier studies in the rat that have suggested the presence of a combinatorial map.
Point-scanning two-photon microscopy enables high-resolution imaging within scattering specimens such as the mammalian brain, but sequential acquisition of voxels fundamentally limits imaging speed. We developed a two-photon imaging technique that scans lines of excitation across a focal plane at multiple angles and uses prior information to recover high-resolution images at over 1.4 billion voxels per second. Using a structural image as a prior for recording neural activity, we imaged visually-evoked and spontaneous glutamate release across hundreds of dendritic spines in mice at depths over 250 microns and frame-rates over 1 kHz. Dendritic glutamate transients in anaesthetized mice are synchronized within spatially-contiguous domains spanning tens of microns at frequencies ranging from 1-100 Hz. We demonstrate high-speed recording of acetylcholine and calcium sensors, 3D single-particle tracking, and imaging in densely-labeled cortex. Our method surpasses limits on the speed of raster-scanned imaging imposed by fluorescence lifetime.
Rodents move their whiskers to locate and identify objects. Cortical areas involved in vibrissal somatosensation and sensorimotor integration include the vibrissal area of the primary motor cortex (vM1), primary somatosensory cortex (vS1; barrel cortex), and secondary somatosensory cortex (S2). We mapped local excitatory pathways in each area across all cortical layers using glutamate uncaging and laser scanning photostimulation. We analyzed these maps to derive laminar connectivity matrices describing the average strengths of pathways between individual neurons in different layers and between entire cortical layers. In vM1, the strongest projection was L2/3→L5. In vS1, strong projections were L2/3→L5 and L4→L3. L6 input and output were weak in both areas. In S2, L2/3→L5 exceeded the strength of the ascending L4→L3 projection, and local input to L6 was prominent. The most conserved pathways were L2/3→L5, and the most variable were L4→L2/3 and pathways involving L6. Local excitatory circuits in different cortical areas are organized around a prominent descending pathway from L2/3→L5, suggesting that sensory cortices are elaborations on a basic motor cortex-like plan.