Filter
Associated Lab
- Aguilera Castrejon Lab (2) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (55) Apply Ahrens Lab filter
- Aso Lab (40) Apply Aso Lab filter
- Baker Lab (19) Apply Baker Lab filter
- Betzig Lab (101) Apply Betzig Lab filter
- Beyene Lab (9) Apply Beyene Lab filter
- Bock Lab (14) Apply Bock Lab filter
- Branson Lab (51) Apply Branson Lab filter
- Card Lab (37) Apply Card Lab filter
- Cardona Lab (45) Apply Cardona Lab filter
- Chklovskii Lab (10) Apply Chklovskii Lab filter
- Clapham Lab (14) Apply Clapham Lab filter
- Cui Lab (19) Apply Cui Lab filter
- Darshan Lab (8) Apply Darshan Lab filter
- Dickson Lab (32) Apply Dickson Lab filter
- Druckmann Lab (21) Apply Druckmann Lab filter
- Dudman Lab (38) Apply Dudman Lab filter
- Eddy/Rivas Lab (30) Apply Eddy/Rivas Lab filter
- Egnor Lab (4) Apply Egnor Lab filter
- Espinosa Medina Lab (15) Apply Espinosa Medina Lab filter
- Feliciano Lab (8) Apply Feliciano Lab filter
- Fetter Lab (31) Apply Fetter Lab filter
- FIB-SEM Technology (1) Apply FIB-SEM Technology filter
- Fitzgerald Lab (16) Apply Fitzgerald Lab filter
- Freeman Lab (15) Apply Freeman Lab filter
- Funke Lab (39) Apply Funke Lab filter
- Gonen Lab (59) Apply Gonen Lab filter
- Grigorieff Lab (34) Apply Grigorieff Lab filter
- Harris Lab (53) Apply Harris Lab filter
- Heberlein Lab (13) Apply Heberlein Lab filter
- Hermundstad Lab (24) Apply Hermundstad Lab filter
- Hess Lab (74) Apply Hess Lab filter
- Ilanges Lab (2) Apply Ilanges Lab filter
- Jayaraman Lab (42) Apply Jayaraman Lab filter
- Ji Lab (33) Apply Ji Lab filter
- Johnson Lab (1) Apply Johnson Lab filter
- Karpova Lab (13) Apply Karpova Lab filter
- Keleman Lab (8) Apply Keleman Lab filter
- Keller Lab (61) Apply Keller Lab filter
- Koay Lab (2) Apply Koay Lab filter
- Lavis Lab (139) Apply Lavis Lab filter
- Lee (Albert) Lab (29) Apply Lee (Albert) Lab filter
- Leonardo Lab (19) Apply Leonardo Lab filter
- Li Lab (4) Apply Li Lab filter
- Lippincott-Schwartz Lab (100) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (2) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (59) Apply Liu (Zhe) Lab filter
- Looger Lab (137) Apply Looger Lab filter
- Magee Lab (31) Apply Magee Lab filter
- Menon Lab (12) Apply Menon Lab filter
- Murphy Lab (6) Apply Murphy Lab filter
- O'Shea Lab (6) Apply O'Shea Lab filter
- Otopalik Lab (1) Apply Otopalik Lab filter
- Pachitariu Lab (36) Apply Pachitariu Lab filter
- Pastalkova Lab (5) Apply Pastalkova Lab filter
- Pavlopoulos Lab (7) Apply Pavlopoulos Lab filter
- Pedram Lab (4) Apply Pedram Lab filter
- Podgorski Lab (16) Apply Podgorski Lab filter
- Reiser Lab (45) Apply Reiser Lab filter
- Riddiford Lab (20) Apply Riddiford Lab filter
- Romani Lab (31) Apply Romani Lab filter
- Rubin Lab (107) Apply Rubin Lab filter
- Saalfeld Lab (46) Apply Saalfeld Lab filter
- Satou Lab (1) Apply Satou Lab filter
- Scheffer Lab (36) Apply Scheffer Lab filter
- Schreiter Lab (51) Apply Schreiter Lab filter
- Sgro Lab (1) Apply Sgro Lab filter
- Shroff Lab (31) Apply Shroff Lab filter
- Simpson Lab (18) Apply Simpson Lab filter
- Singer Lab (37) Apply Singer Lab filter
- Spruston Lab (58) Apply Spruston Lab filter
- Stern Lab (73) Apply Stern Lab filter
- Sternson Lab (47) Apply Sternson Lab filter
- Stringer Lab (33) Apply Stringer Lab filter
- Svoboda Lab (131) Apply Svoboda Lab filter
- Tebo Lab (9) Apply Tebo Lab filter
- Tervo Lab (9) Apply Tervo Lab filter
- Tillberg Lab (18) Apply Tillberg Lab filter
- Tjian Lab (17) Apply Tjian Lab filter
- Truman Lab (58) Apply Truman Lab filter
- Turaga Lab (40) Apply Turaga Lab filter
- Turner Lab (28) Apply Turner Lab filter
- Vale Lab (8) Apply Vale Lab filter
- Voigts Lab (3) Apply Voigts Lab filter
- Wang (Meng) Lab (22) Apply Wang (Meng) Lab filter
- Wang (Shaohe) Lab (6) Apply Wang (Shaohe) Lab filter
- Wu Lab (8) Apply Wu Lab filter
- Zlatic Lab (26) Apply Zlatic Lab filter
- Zuker Lab (5) Apply Zuker Lab filter
Associated Project Team
- CellMap (12) Apply CellMap filter
- COSEM (3) Apply COSEM filter
- FIB-SEM Technology (3) Apply FIB-SEM Technology filter
- Fly Descending Interneuron (11) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (14) Apply Fly Functional Connectome filter
- Fly Olympiad (5) Apply Fly Olympiad filter
- FlyEM (54) Apply FlyEM filter
- FlyLight (49) Apply FlyLight filter
- GENIE (47) Apply GENIE filter
- Integrative Imaging (6) Apply Integrative Imaging filter
- Larval Olympiad (2) Apply Larval Olympiad filter
- MouseLight (18) Apply MouseLight filter
- NeuroSeq (1) Apply NeuroSeq filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (27) Apply Tool Translation Team (T3) filter
- Transcription Imaging (45) Apply Transcription Imaging filter
Associated Support Team
- Project Pipeline Support (5) Apply Project Pipeline Support filter
- Anatomy and Histology (18) Apply Anatomy and Histology filter
- Cryo-Electron Microscopy (39) Apply Cryo-Electron Microscopy filter
- Electron Microscopy (17) Apply Electron Microscopy filter
- Gene Targeting and Transgenics (11) Apply Gene Targeting and Transgenics filter
- High Performance Computing (7) Apply High Performance Computing filter
- Integrative Imaging (17) Apply Integrative Imaging filter
- Invertebrate Shared Resource (40) Apply Invertebrate Shared Resource filter
- Janelia Experimental Technology (37) Apply Janelia Experimental Technology filter
- Management Team (1) Apply Management Team filter
- Molecular Genomics (15) Apply Molecular Genomics filter
- Primary & iPS Cell Culture (14) Apply Primary & iPS Cell Culture filter
- Project Technical Resources (50) Apply Project Technical Resources filter
- Quantitative Genomics (19) Apply Quantitative Genomics filter
- Scientific Computing (93) Apply Scientific Computing filter
- Viral Tools (14) Apply Viral Tools filter
- Vivarium (7) Apply Vivarium filter
Publication Date
- 2025 (160) Apply 2025 filter
- 2024 (213) Apply 2024 filter
- 2023 (158) Apply 2023 filter
- 2022 (166) Apply 2022 filter
- 2021 (175) Apply 2021 filter
- 2020 (177) Apply 2020 filter
- 2019 (177) Apply 2019 filter
- 2018 (206) Apply 2018 filter
- 2017 (186) Apply 2017 filter
- 2016 (191) Apply 2016 filter
- 2015 (195) Apply 2015 filter
- 2014 (190) Apply 2014 filter
- 2013 (136) Apply 2013 filter
- 2012 (112) Apply 2012 filter
- 2011 (98) Apply 2011 filter
- 2010 (61) Apply 2010 filter
- 2009 (56) Apply 2009 filter
- 2008 (40) Apply 2008 filter
- 2007 (21) Apply 2007 filter
- 2006 (3) Apply 2006 filter
2721 Janelia Publications
Showing 2081-2090 of 2721 resultsThe Rfam database aims to catalogue non-coding RNAs through the use of sequence alignments and statistical profile models known as covariance models. In this contribution, we discuss the pros and cons of using the online encyclopedia, Wikipedia, as a source of community-derived annotation. We discuss the addition of groupings of related RNA families into clans and new developments to the website. Rfam is available on the Web at http://rfam.sanger.ac.uk.
Enzymes that cut proteins inside membranes regulate diverse cellular events, including cell signaling, homeostasis, and host-pathogen interactions. Adaptations that enable catalysis in this exceptional environment are poorly understood. We visualized single molecules of multiple rhomboid intramembrane proteases and unrelated proteins in living cells (human and ) and planar lipid bilayers. Notably, only rhomboid proteins were able to diffuse above the Saffman-Delbrück viscosity limit of the membrane. Hydrophobic mismatch with the irregularly shaped rhomboid fold distorted surrounding lipids and propelled rhomboid diffusion. The rate of substrate processing in living cells scaled with rhomboid diffusivity. Thus, intramembrane proteolysis is naturally diffusion-limited, but cells mitigate this constraint by using the rhomboid fold to overcome the "speed limit" of membrane diffusion.
Stringent response is a conserved bacterial stress response underlying virulence and antibiotic resistance. RelA/SpoT-homolog proteins synthesize transcriptional modulators (p)ppGpp, allowing bacteria to adapt to stress. RelA is activated during amino-acid starvation, when cognate deacyl-tRNA binds to the ribosomal A (aminoacyl-tRNA) site. We report four cryo-EM structures of E. coli RelA bound to the 70S ribosome, in the absence and presence of deacyl-tRNA accommodating in the 30S A site. The boomerang-shaped RelA with a wingspan of more than 100 Å wraps around the A/R (30S A-site/RelA-bound) tRNA. The CCA end of the A/R tRNA pins the central TGS domain against the 30S subunit, presenting the (p)ppGpp-synthetase domain near the 30S spur. The ribosome and A/R tRNA are captured in three conformations, revealing hitherto elusive states of tRNA engagement with the ribosomal decoding center. Decoding-center rearrangements are coupled with the step-wise 30S-subunit 'closure', providing insights into the dynamics of high-fidelity tRNA decoding.
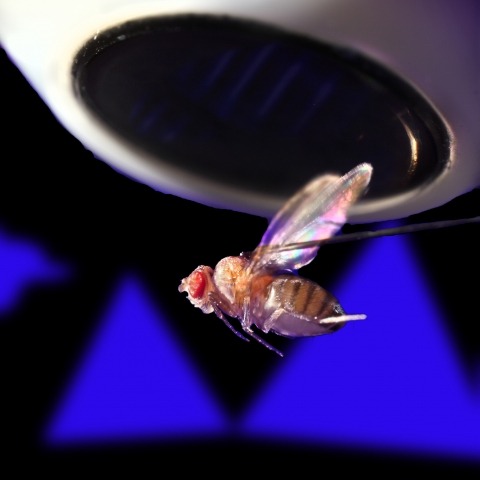
Ring attractors are a class of recurrent networks hypothesized to underlie the representation of heading direction. Such network structures, schematized as a ring of neurons whose connectivity depends on their heading preferences, can sustain a bump-like activity pattern whose location can be updated by continuous shifts along either turn direction. We recently reported that a population of fly neurons represents the animal's heading via bump-like activity dynamics. We combined two-photon calcium imaging in head-fixed flying flies with optogenetics to overwrite the existing population representation with an artificial one, which was then maintained by the circuit with naturalistic dynamics. A network with local excitation and global inhibition enforces this unique and persistent heading representation. Ring attractor networks have long been invoked in theoretical work; our study provides physiological evidence of their existence and functional architecture.

A number of recent studies have provided compelling demonstrations that both mice and rats can be trained to perform a variety of behavioral tasks while restrained by mechanical elements mounted to the skull. The independent development of this technique by a number of laboratories has led to diverse solutions. We found that these solutions often used expensive materials and impeded future development and modification in the absence of engineering support. In order to address these issues, here we report on the development of a flexible single hardware design for electrophysiology and imaging both in brain tissue in vitro. Our hardware facilitates the rapid conversion of a single preparation between physiology and imaging system and the conversion of a given system between preparations. In addition, our use of rapid prototyping machines ("3D printers") allows for the deployment of new designs within a day. Here, we present specifications for design and manufacturing as well as some data from our lab demonstrating the suitability of the design for physiology in behaving animals and imaging in vitro and in vivo.
Long-distance RNA transport enables local protein synthesis at metabolically-active sites distant from the nucleus. This process ensures an appropriate spatial organization of proteins, vital to polarized cells such as neurons. Here, we present a mechanism for RNA transport in which RNA granules "hitchhike" on moving lysosomes. In vitro biophysical modeling, live-cell microscopy, and unbiased proximity labeling proteomics reveal that annexin A11 (ANXA11), an RNA granule-associated phosphoinositide-binding protein, acts as a molecular tether between RNA granules and lysosomes. ANXA11 possesses an N-terminal low complexity domain, facilitating its phase separation into membraneless RNA granules, and a C-terminal membrane binding domain, enabling interactions with lysosomes. RNA granule transport requires ANXA11, and amyotrophic lateral sclerosis (ALS)-associated mutations in ANXA11 impair RNA granule transport by disrupting their interactions with lysosomes. Thus, ANXA11 mediates neuronal RNA transport by tethering RNA granules to actively-transported lysosomes, performing a critical cellular function that is disrupted in ALS.
Protein clustering is a hallmark of genome regulation in mammalian cells. However, the dynamic molecular processes involved make it difficult to correlate clustering with functional consequences in vivo. We developed a live-cell super-resolution approach to uncover the correlation between mRNA synthesis and the dynamics of RNA Polymerase II (Pol II) clusters at a gene locus. For endogenous β-actin genes in mouse embryonic fibroblasts, we observe that short-lived (~8 s) Pol II clusters correlate with basal mRNA output. During serum stimulation, a stereotyped increase in Pol II cluster lifetime correlates with a proportionate increase in the number of mRNAs synthesized. Our findings suggest that transient clustering of Pol II may constitute a pre-transcriptional regulatory event that predictably modulates nascent mRNA output.
Neurons decentralize protein synthesis from the cell body to support the active metabolism of remote dendritic and axonal compartments. The neuronal RNA transport apparatus, composed of cis-acting RNA regulatory elements, neuronal transport granule proteins, and motor adaptor complexes, drives the long-distance RNA trafficking required for local protein synthesis. Over the past decade, advances in human genetics, subcellular biochemistry, and high-resolution imaging have implicated each member of the apparatus in several neurodegenerative diseases, establishing failed RNA transport and associated processes as a unifying pathomechanism. In this review, we deconstruct the RNA transport apparatus, exploring each constituent's role in RNA localization and illuminating their unique contributions to neurodegeneration.
Localized protein translation is critical in many biological contexts, particularly in highly polarized cells, such as neurons, to regulate gene expression in a spatiotemporal manner. The cytoplasmic polyadenylation element-binding (CPEB) family of RNA-binding proteins has emerged as a key regulator of mRNA transport and local translation required for early embryonic development, synaptic plasticity, and long-term memory (LTM). Drosophila Orb and Orb2 are single members of the CPEB1 and CPEB2 subfamilies of the CPEB proteins, respectively. At present, the identity of the mRNA targets they regulate is not fully known, and the binding specificity of the CPEB2 subfamily is a matter of debate. Using transcriptome-wide UV cross-linking and immunoprecipitation, we define the mRNA-binding sites and targets of Drosophila CPEBs. Both Orb and Orb2 bind linear cytoplasmic polyadenylation element-like sequences in the 3' UTRs of largely overlapping target mRNAs, with Orb2 potentially having a broader specificity. Both proteins use their RNA-recognition motifs but not the Zinc-finger region for RNA binding. A subset of Orb2 targets is translationally regulated in cultured S2 cells and fly head extracts. Moreover, pan-neuronal RNAi knockdown of these targets suggests that a number of these targets are involved in LTM. Our results provide a comprehensive list of mRNA targets of the two CPEB proteins in Drosophila, thus providing insights into local protein synthesis involved in various biological processes, including LTM.
Bacterial Rho-independent terminators (RITs) are important genomic landmarks involved in gene regulation and terminating gene expression. In this investigation we present RNIE, a probabilistic approach for predicting RITs. The method is based upon covariance models which have been known for many years to be the most accurate computational tools for predicting homology in structural non-coding RNAs. We show that RNIE has superior performance in model species from a spectrum of bacterial phyla. Further analysis of species where a low number of RITs were predicted revealed a highly conserved structural sequence motif enriched near the genic termini of the pathogenic Actinobacteria, Mycobacterium tuberculosis. This motif, together with classical RITs, account for up to 90% of all the significantly structured regions from the termini of M. tuberculosis genic elements. The software, predictions and alignments described below are available from http://github.com/ppgardne/RNIE.