Filter
Associated Lab
- Aguilera Castrejon Lab (1) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (53) Apply Ahrens Lab filter
- Aso Lab (40) Apply Aso Lab filter
- Baker Lab (19) Apply Baker Lab filter
- Betzig Lab (101) Apply Betzig Lab filter
- Beyene Lab (8) Apply Beyene Lab filter
- Bock Lab (14) Apply Bock Lab filter
- Branson Lab (49) Apply Branson Lab filter
- Card Lab (36) Apply Card Lab filter
- Cardona Lab (45) Apply Cardona Lab filter
- Chklovskii Lab (10) Apply Chklovskii Lab filter
- Clapham Lab (14) Apply Clapham Lab filter
- Cui Lab (19) Apply Cui Lab filter
- Darshan Lab (8) Apply Darshan Lab filter
- Dickson Lab (32) Apply Dickson Lab filter
- Druckmann Lab (21) Apply Druckmann Lab filter
- Dudman Lab (38) Apply Dudman Lab filter
- Eddy/Rivas Lab (30) Apply Eddy/Rivas Lab filter
- Egnor Lab (4) Apply Egnor Lab filter
- Espinosa Medina Lab (15) Apply Espinosa Medina Lab filter
- Feliciano Lab (7) Apply Feliciano Lab filter
- Fetter Lab (31) Apply Fetter Lab filter
- Fitzgerald Lab (16) Apply Fitzgerald Lab filter
- Freeman Lab (15) Apply Freeman Lab filter
- Funke Lab (38) Apply Funke Lab filter
- Gonen Lab (59) Apply Gonen Lab filter
- Grigorieff Lab (34) Apply Grigorieff Lab filter
- Harris Lab (53) Apply Harris Lab filter
- Heberlein Lab (13) Apply Heberlein Lab filter
- Hermundstad Lab (22) Apply Hermundstad Lab filter
- Hess Lab (74) Apply Hess Lab filter
- Ilanges Lab (2) Apply Ilanges Lab filter
- Jayaraman Lab (42) Apply Jayaraman Lab filter
- Ji Lab (33) Apply Ji Lab filter
- Johnson Lab (1) Apply Johnson Lab filter
- Karpova Lab (13) Apply Karpova Lab filter
- Keleman Lab (8) Apply Keleman Lab filter
- Keller Lab (61) Apply Keller Lab filter
- Koay Lab (2) Apply Koay Lab filter
- Lavis Lab (137) Apply Lavis Lab filter
- Lee (Albert) Lab (29) Apply Lee (Albert) Lab filter
- Leonardo Lab (19) Apply Leonardo Lab filter
- Li Lab (4) Apply Li Lab filter
- Lippincott-Schwartz Lab (97) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (1) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (58) Apply Liu (Zhe) Lab filter
- Looger Lab (137) Apply Looger Lab filter
- Magee Lab (31) Apply Magee Lab filter
- Menon Lab (12) Apply Menon Lab filter
- Murphy Lab (6) Apply Murphy Lab filter
- O'Shea Lab (6) Apply O'Shea Lab filter
- Otopalik Lab (1) Apply Otopalik Lab filter
- Pachitariu Lab (36) Apply Pachitariu Lab filter
- Pastalkova Lab (5) Apply Pastalkova Lab filter
- Pavlopoulos Lab (7) Apply Pavlopoulos Lab filter
- Pedram Lab (4) Apply Pedram Lab filter
- Podgorski Lab (16) Apply Podgorski Lab filter
- Reiser Lab (45) Apply Reiser Lab filter
- Riddiford Lab (20) Apply Riddiford Lab filter
- Romani Lab (31) Apply Romani Lab filter
- Rubin Lab (105) Apply Rubin Lab filter
- Saalfeld Lab (46) Apply Saalfeld Lab filter
- Satou Lab (1) Apply Satou Lab filter
- Scheffer Lab (36) Apply Scheffer Lab filter
- Schreiter Lab (50) Apply Schreiter Lab filter
- Sgro Lab (1) Apply Sgro Lab filter
- Shroff Lab (31) Apply Shroff Lab filter
- Simpson Lab (18) Apply Simpson Lab filter
- Singer Lab (37) Apply Singer Lab filter
- Spruston Lab (57) Apply Spruston Lab filter
- Stern Lab (73) Apply Stern Lab filter
- Sternson Lab (47) Apply Sternson Lab filter
- Stringer Lab (32) Apply Stringer Lab filter
- Svoboda Lab (131) Apply Svoboda Lab filter
- Tebo Lab (9) Apply Tebo Lab filter
- Tervo Lab (9) Apply Tervo Lab filter
- Tillberg Lab (18) Apply Tillberg Lab filter
- Tjian Lab (17) Apply Tjian Lab filter
- Truman Lab (58) Apply Truman Lab filter
- Turaga Lab (39) Apply Turaga Lab filter
- Turner Lab (27) Apply Turner Lab filter
- Vale Lab (7) Apply Vale Lab filter
- Voigts Lab (3) Apply Voigts Lab filter
- Wang (Meng) Lab (21) Apply Wang (Meng) Lab filter
- Wang (Shaohe) Lab (6) Apply Wang (Shaohe) Lab filter
- Wu Lab (8) Apply Wu Lab filter
- Zlatic Lab (26) Apply Zlatic Lab filter
- Zuker Lab (5) Apply Zuker Lab filter
Associated Project Team
- CellMap (12) Apply CellMap filter
- COSEM (3) Apply COSEM filter
- FIB-SEM Technology (3) Apply FIB-SEM Technology filter
- Fly Descending Interneuron (11) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (14) Apply Fly Functional Connectome filter
- Fly Olympiad (5) Apply Fly Olympiad filter
- FlyEM (53) Apply FlyEM filter
- FlyLight (49) Apply FlyLight filter
- GENIE (46) Apply GENIE filter
- Integrative Imaging (4) Apply Integrative Imaging filter
- Larval Olympiad (2) Apply Larval Olympiad filter
- MouseLight (18) Apply MouseLight filter
- NeuroSeq (1) Apply NeuroSeq filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (26) Apply Tool Translation Team (T3) filter
- Transcription Imaging (45) Apply Transcription Imaging filter
Associated Support Team
- Project Pipeline Support (5) Apply Project Pipeline Support filter
- Anatomy and Histology (18) Apply Anatomy and Histology filter
- Cryo-Electron Microscopy (34) Apply Cryo-Electron Microscopy filter
- Electron Microscopy (15) Apply Electron Microscopy filter
- Gene Targeting and Transgenics (11) Apply Gene Targeting and Transgenics filter
- Integrative Imaging (17) Apply Integrative Imaging filter
- Invertebrate Shared Resource (40) Apply Invertebrate Shared Resource filter
- Janelia Experimental Technology (37) Apply Janelia Experimental Technology filter
- Management Team (1) Apply Management Team filter
- Molecular Genomics (15) Apply Molecular Genomics filter
- Primary & iPS Cell Culture (14) Apply Primary & iPS Cell Culture filter
- Project Technical Resources (50) Apply Project Technical Resources filter
- Quantitative Genomics (19) Apply Quantitative Genomics filter
- Scientific Computing Software (92) Apply Scientific Computing Software filter
- Scientific Computing Systems (7) Apply Scientific Computing Systems filter
- Viral Tools (14) Apply Viral Tools filter
- Vivarium (7) Apply Vivarium filter
Publication Date
- 2025 (119) Apply 2025 filter
- 2024 (216) Apply 2024 filter
- 2023 (159) Apply 2023 filter
- 2022 (167) Apply 2022 filter
- 2021 (175) Apply 2021 filter
- 2020 (177) Apply 2020 filter
- 2019 (177) Apply 2019 filter
- 2018 (206) Apply 2018 filter
- 2017 (186) Apply 2017 filter
- 2016 (191) Apply 2016 filter
- 2015 (195) Apply 2015 filter
- 2014 (190) Apply 2014 filter
- 2013 (136) Apply 2013 filter
- 2012 (112) Apply 2012 filter
- 2011 (98) Apply 2011 filter
- 2010 (61) Apply 2010 filter
- 2009 (56) Apply 2009 filter
- 2008 (40) Apply 2008 filter
- 2007 (21) Apply 2007 filter
- 2006 (3) Apply 2006 filter
2685 Janelia Publications
Showing 2161-2170 of 2685 resultsThe hippocampal CA3 region is essential for pattern completion and generation of sharp-wave ripples. During these operations, coordinated activation of ensembles of CA3 pyramidal neurons produces spatiotemporally structured input patterns arriving onto dendrites of recurrently connected CA3 neurons. To understand how such input patterns are translated into specific output patterns, we characterized dendritic integration in CA3 pyramidal cells using two-photon imaging and glutamate uncaging. We found that thin dendrites of CA3 pyramidal neurons integrate synchronous synaptic input in a highly supralinear fashion. The amplification was primarily mediated by NMDA receptor activation and was present over a relatively broad range of spatiotemporal input patterns. The decay of voltage responses, temporal summation, and action potential output was regulated in a compartmentalized fashion mainly by a G-protein-activated inwardly rectifying K(+) current. Our results suggest that plastic dendritic integrative mechanisms may support ensemble behavior in pyramidal neurons of the hippocampal circuitry.
The gills of most teleost fishes are covered by plate-like structures, the secondary lamellae, that provide the bulk of the respiratory surface area. Water passing over the secondary lamellae exchanges gases with blood passing through the secondary lamellae, forming a system that has served as a classic model of counter-current exchange. In this study, a computational model of flow around the secondary lamellae is used to examine the hydrodynamic consequences of changes to the lamellar morphology. Consistent with previous studies, the interlamellar distance is found to strongly affect the hydrodynamic resistance of the gills. However, the presence of a small gap between the tips of the secondary lamellae is found to have a similarly strong effect on the hydrodynamic resistance and flow patterns within the gills. The results from this model have been generally formulated, allowing the calculation of the hydrodynamic resistance for measured morphometric parameters. These results provide a new basis for comparing theoretical predictions of the gill resistance with measured values, and provide a general model for examining the diversity gill morphologies observed in teleost fishes.
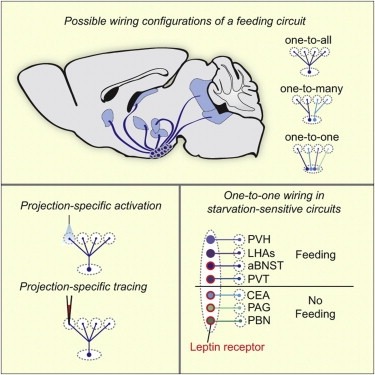
Neural circuits for essential natural behaviors are shaped by selective pressure to coordinate reliable execution of flexible goal-directed actions. However, the structural and functional organization of survival-oriented circuits is poorly understood due to exceptionally complex neuroanatomy. This is exemplified by AGRP neurons, which are a molecularly defined population that is sufficient to rapidly coordinate voracious food seeking and consumption behaviors. Here, we use cell-type-specific techniques for neural circuit manipulation and projection-specific anatomical analysis to examine the organization of this critical homeostatic circuit that regulates feeding. We show that AGRP neuronal circuits use a segregated, parallel, and redundant output configuration. AGRP neuron axon projections that target different brain regions originate from distinct subpopulations, several of which are sufficient to independently evoke feeding. The concerted anatomical and functional analysis of AGRP neuron projection populations reveals a constellation of core forebrain nodes, which are part of an extended circuit that mediates feeding behavior.
Mapping mammalian synaptic connectivity has long been an important goal of neuroscientists since it is considered crucial for explaining human perception and behavior. Yet, despite enormous efforts, the overwhelming complexity of the neural circuitry and the lack of appropriate techniques to unravel it have limited the success of efforts to map connectivity. However, recent technological advances designed to overcome the limitations of conventional methods for connectivity mapping may bring about a turning point. Here, we address the promises and pitfalls of these new mapping technologies.
A new generation of direct electron detectors for transmission electron microscopy (TEM) promises significant improvement over previous detectors in terms of their modulation transfer function (MTF) and detective quantum efficiency (DQE). However, the performance of these new detectors needs to be carefully monitored in order to optimize imaging conditions and check for degradation over time. We have developed an easy-to-use software tool, FindDQE, to measure MTF and DQE of electron detectors using images of a microscope’s built-in beam stop. Using this software, we have determined the DQE curves of four direct electron detectors currently available: the Gatan K2 Summit, the FEI Falcon I and II, and the Direct Electron DE-12, under a variety of total dose and dose rate conditions. We have additionally measured the curves for the Gatan US4000 and TVIPS TemCam-F416 scintillator-based cameras. We compare the results from our new method with published curves.
Many mammals forage and burrow in dark constrained spaces. Touch through facial whiskers is important during these activities, but the close quarters makes whisker deployment challenging. The diverse shapes of facial whiskers reflect distinct ecological niches. Rodent whiskers are conical, often with a remarkably linear taper. Here we use theoretical and experimental methods to analyze interactions of mouse whiskers with objects. When pushed into objects, conical whiskers suddenly slip at a critical angle. In contrast, cylindrical whiskers do not slip for biologically plausible movements. Conical whiskers sweep across objects and textures in characteristic sequences of brief sticks and slips, which provide information about the tactile world. In contrast, cylindrical whiskers stick and remain stuck, even when sweeping across fine textures. Thus the conical whisker structure is adaptive for sensor mobility in constrained environments and in feature extraction during active haptic exploration of objects and surfaces. DOI: http://dx.doi.org/10.7554/eLife.01350.001.
We demonstrate that it is feasible to determine high-resolution protein structures by electron crystallography of three-dimensional crystals in an electron cryo-microscope (CryoEM). Lysozyme microcrystals were frozen on an electron microscopy grid, and electron diffraction data collected to 1.7 Å resolution. We developed a data collection protocol to collect a full-tilt series in electron diffraction to atomic resolution. A single tilt series contains up to 90 individual diffraction patterns collected from a single crystal with tilt angle increment of 0.1–1° and a total accumulated electron dose less than 10 electrons per angstrom squared. We indexed the data from three crystals and used them for structure determination of lysozyme by molecular replacement followed by crystallographic refinement to 2.9 Å resolution. This proof of principle paves the way for the implementation of a new technique, which we name ‘MicroED’, that may have wide applicability in structural biology.
Many animals utilize acoustic signals-or songs-to attract mates. During courtship, Drosophila melanogaster males vibrate a wing to produce trains of pulses and extended tone, called pulse and sine song, respectively. Courtship songs in the genus Drosophila are exceedingly diverse, and different song features appear to have evolved independently of each other. How the nervous system allows such diversity to evolve is not understood. Here, we identify a wing muscle in D. melanogaster (hg1) that is uniquely male-enlarged. The hg1 motoneuron and the sexually dimorphic development of the hg1 muscle are required specifically for the sine component of the male song. In contrast, the motoneuron innervating a sexually monomorphic wing muscle, ps1, is required specifically for a feature of pulse song. Thus, individual wing motor pathways can control separate aspects of courtship song and may provide a "modular" anatomical substrate for the evolution of diverse songs.
The subcellular locations of synapses on pyramidal neurons strongly influences dendritic integration and synaptic plasticity. Despite this, there is little quantitative data on spatial distributions of specific types of synaptic input. Here we use array tomography (AT), a high-resolution optical microscopy method, to examine thalamocortical (TC) input onto layer 5 pyramidal neurons. We first verified the ability of AT to identify synapses using parallel electron microscopic analysis of TC synapses in layer 4. We then use large-scale array tomography (LSAT) to measure TC synapse distribution on L5 pyramidal neurons in a 1.00 × 0.83 × 0.21 mm(3) volume of mouse somatosensory cortex. We found that TC synapses primarily target basal dendrites in layer 5, but also make a considerable input to proximal apical dendrites in L4, consistent with previous work. Our analysis further suggests that TC inputs are biased toward certain branches and, within branches, synapses show significant clustering with an excess of TC synapse nearest neighbors within 5-15 μm compared to a random distribution. Thus, we show that AT is a sensitive and quantitative method to map specific types of synaptic input on the dendrites of entire neurons. We anticipate that this technique will be of wide utility for mapping functionally-relevant anatomical connectivity in neural circuits.