Filter
Associated Lab
- Aguilera Castrejon Lab (1) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (53) Apply Ahrens Lab filter
- Aso Lab (40) Apply Aso Lab filter
- Baker Lab (19) Apply Baker Lab filter
- Betzig Lab (101) Apply Betzig Lab filter
- Beyene Lab (8) Apply Beyene Lab filter
- Bock Lab (14) Apply Bock Lab filter
- Branson Lab (50) Apply Branson Lab filter
- Card Lab (36) Apply Card Lab filter
- Cardona Lab (45) Apply Cardona Lab filter
- Chklovskii Lab (10) Apply Chklovskii Lab filter
- Clapham Lab (14) Apply Clapham Lab filter
- Cui Lab (19) Apply Cui Lab filter
- Darshan Lab (8) Apply Darshan Lab filter
- Dickson Lab (32) Apply Dickson Lab filter
- Druckmann Lab (21) Apply Druckmann Lab filter
- Dudman Lab (38) Apply Dudman Lab filter
- Eddy/Rivas Lab (30) Apply Eddy/Rivas Lab filter
- Egnor Lab (4) Apply Egnor Lab filter
- Espinosa Medina Lab (15) Apply Espinosa Medina Lab filter
- Feliciano Lab (7) Apply Feliciano Lab filter
- Fetter Lab (31) Apply Fetter Lab filter
- Fitzgerald Lab (16) Apply Fitzgerald Lab filter
- Freeman Lab (15) Apply Freeman Lab filter
- Funke Lab (38) Apply Funke Lab filter
- Gonen Lab (59) Apply Gonen Lab filter
- Grigorieff Lab (34) Apply Grigorieff Lab filter
- Harris Lab (53) Apply Harris Lab filter
- Heberlein Lab (13) Apply Heberlein Lab filter
- Hermundstad Lab (22) Apply Hermundstad Lab filter
- Hess Lab (74) Apply Hess Lab filter
- Ilanges Lab (2) Apply Ilanges Lab filter
- Jayaraman Lab (42) Apply Jayaraman Lab filter
- Ji Lab (33) Apply Ji Lab filter
- Johnson Lab (1) Apply Johnson Lab filter
- Karpova Lab (13) Apply Karpova Lab filter
- Keleman Lab (8) Apply Keleman Lab filter
- Keller Lab (61) Apply Keller Lab filter
- Koay Lab (2) Apply Koay Lab filter
- Lavis Lab (137) Apply Lavis Lab filter
- Lee (Albert) Lab (29) Apply Lee (Albert) Lab filter
- Leonardo Lab (19) Apply Leonardo Lab filter
- Li Lab (4) Apply Li Lab filter
- Lippincott-Schwartz Lab (97) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (1) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (58) Apply Liu (Zhe) Lab filter
- Looger Lab (137) Apply Looger Lab filter
- Magee Lab (31) Apply Magee Lab filter
- Menon Lab (12) Apply Menon Lab filter
- Murphy Lab (6) Apply Murphy Lab filter
- O'Shea Lab (6) Apply O'Shea Lab filter
- Otopalik Lab (1) Apply Otopalik Lab filter
- Pachitariu Lab (36) Apply Pachitariu Lab filter
- Pastalkova Lab (5) Apply Pastalkova Lab filter
- Pavlopoulos Lab (7) Apply Pavlopoulos Lab filter
- Pedram Lab (4) Apply Pedram Lab filter
- Podgorski Lab (16) Apply Podgorski Lab filter
- Reiser Lab (45) Apply Reiser Lab filter
- Riddiford Lab (20) Apply Riddiford Lab filter
- Romani Lab (31) Apply Romani Lab filter
- Rubin Lab (105) Apply Rubin Lab filter
- Saalfeld Lab (46) Apply Saalfeld Lab filter
- Satou Lab (1) Apply Satou Lab filter
- Scheffer Lab (36) Apply Scheffer Lab filter
- Schreiter Lab (50) Apply Schreiter Lab filter
- Sgro Lab (1) Apply Sgro Lab filter
- Shroff Lab (31) Apply Shroff Lab filter
- Simpson Lab (18) Apply Simpson Lab filter
- Singer Lab (37) Apply Singer Lab filter
- Spruston Lab (57) Apply Spruston Lab filter
- Stern Lab (73) Apply Stern Lab filter
- Sternson Lab (47) Apply Sternson Lab filter
- Stringer Lab (32) Apply Stringer Lab filter
- Svoboda Lab (131) Apply Svoboda Lab filter
- Tebo Lab (9) Apply Tebo Lab filter
- Tervo Lab (9) Apply Tervo Lab filter
- Tillberg Lab (18) Apply Tillberg Lab filter
- Tjian Lab (17) Apply Tjian Lab filter
- Truman Lab (58) Apply Truman Lab filter
- Turaga Lab (39) Apply Turaga Lab filter
- Turner Lab (27) Apply Turner Lab filter
- Vale Lab (7) Apply Vale Lab filter
- Voigts Lab (3) Apply Voigts Lab filter
- Wang (Meng) Lab (21) Apply Wang (Meng) Lab filter
- Wang (Shaohe) Lab (6) Apply Wang (Shaohe) Lab filter
- Wu Lab (8) Apply Wu Lab filter
- Zlatic Lab (26) Apply Zlatic Lab filter
- Zuker Lab (5) Apply Zuker Lab filter
Associated Project Team
- CellMap (12) Apply CellMap filter
- COSEM (3) Apply COSEM filter
- FIB-SEM Technology (3) Apply FIB-SEM Technology filter
- Fly Descending Interneuron (11) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (14) Apply Fly Functional Connectome filter
- Fly Olympiad (5) Apply Fly Olympiad filter
- FlyEM (53) Apply FlyEM filter
- FlyLight (49) Apply FlyLight filter
- GENIE (46) Apply GENIE filter
- Integrative Imaging (4) Apply Integrative Imaging filter
- Larval Olympiad (2) Apply Larval Olympiad filter
- MouseLight (18) Apply MouseLight filter
- NeuroSeq (1) Apply NeuroSeq filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (26) Apply Tool Translation Team (T3) filter
- Transcription Imaging (45) Apply Transcription Imaging filter
Associated Support Team
- Project Pipeline Support (5) Apply Project Pipeline Support filter
- Anatomy and Histology (18) Apply Anatomy and Histology filter
- Cryo-Electron Microscopy (35) Apply Cryo-Electron Microscopy filter
- Electron Microscopy (15) Apply Electron Microscopy filter
- Gene Targeting and Transgenics (11) Apply Gene Targeting and Transgenics filter
- Integrative Imaging (17) Apply Integrative Imaging filter
- Invertebrate Shared Resource (40) Apply Invertebrate Shared Resource filter
- Janelia Experimental Technology (37) Apply Janelia Experimental Technology filter
- Management Team (1) Apply Management Team filter
- Molecular Genomics (15) Apply Molecular Genomics filter
- Primary & iPS Cell Culture (14) Apply Primary & iPS Cell Culture filter
- Project Technical Resources (50) Apply Project Technical Resources filter
- Quantitative Genomics (19) Apply Quantitative Genomics filter
- Scientific Computing Software (92) Apply Scientific Computing Software filter
- Scientific Computing Systems (7) Apply Scientific Computing Systems filter
- Viral Tools (14) Apply Viral Tools filter
- Vivarium (7) Apply Vivarium filter
Publication Date
- 2025 (122) Apply 2025 filter
- 2024 (215) Apply 2024 filter
- 2023 (159) Apply 2023 filter
- 2022 (167) Apply 2022 filter
- 2021 (175) Apply 2021 filter
- 2020 (177) Apply 2020 filter
- 2019 (177) Apply 2019 filter
- 2018 (206) Apply 2018 filter
- 2017 (186) Apply 2017 filter
- 2016 (191) Apply 2016 filter
- 2015 (195) Apply 2015 filter
- 2014 (190) Apply 2014 filter
- 2013 (136) Apply 2013 filter
- 2012 (112) Apply 2012 filter
- 2011 (98) Apply 2011 filter
- 2010 (61) Apply 2010 filter
- 2009 (56) Apply 2009 filter
- 2008 (40) Apply 2008 filter
- 2007 (21) Apply 2007 filter
- 2006 (3) Apply 2006 filter
2687 Janelia Publications
Showing 2221-2230 of 2687 resultsCalmodulin (CaM) is a universal regulatory protein that communicates the presence of calcium to its molecular targets and correspondingly modulates their function. This key signaling protein is important for controlling the activity of hundreds of membrane channels and transporters. However, understanding of the structural mechanisms driving CaM regulation of full-length membrane proteins has remained elusive. In this study, we determined the pseudoatomic structure of full-length mammalian aquaporin-0 (AQP0, Bos taurus) in complex with CaM, using EM to elucidate how this signaling protein modulates water-channel function. Molecular dynamics and functional mutation studies reveal how CaM binding inhibits AQP0 water permeability by allosterically closing the cytoplasmic gate of AQP0. Our mechanistic model provides new insight, only possible in the context of the fully assembled channel, into how CaM regulates multimeric channels by facilitating cooperativity between adjacent subunits.
Fluorescent calcium sensors are widely used to image neural activity. Using structure-based mutagenesis and neuron-based screening, we developed a family of ultrasensitive protein calcium sensors (GCaMP6) that outperformed other sensors in cultured neurons and in zebrafish, flies and mice in vivo. In layer 2/3 pyramidal neurons of the mouse visual cortex, GCaMP6 reliably detected single action potentials in neuronal somata and orientation-tuned synaptic calcium transients in individual dendritic spines. The orientation tuning of structurally persistent spines was largely stable over timescales of weeks. Orientation tuning averaged across spine populations predicted the tuning of their parent cell. Although the somata of GABAergic neurons showed little orientation tuning, their dendrites included highly tuned dendritic segments (5–40-µm long). GCaMP6 sensors thus provide new windows into the organization and dynamics of neural circuits over multiple spatial and temporal scales.
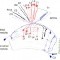
Motion detection is a fundamental neural computation performed by many sensory systems. In the fly, local motion computation is thought to occur within the first two layers of the visual system, the lamina and medulla. We constructed specific genetic driver lines for each of the 12 neuron classes in the lamina. We then depolarized and hyperpolarized each neuron type and quantified fly behavioral responses to a diverse set of motion stimuli. We found that only a small number of lamina output neurons are essential for motion detection, while most neurons serve to sculpt and enhance these feedforward pathways. Two classes of feedback neurons (C2 and C3), and lamina output neurons (L2 and L4), are required for normal detection of directional motion stimuli. Our results reveal a prominent role for feedback and lateral interactions in motion processing and demonstrate that motion-dependent behaviors rely on contributions from nearly all lamina neuron classes.
Genetically hard-wired neural mechanisms must enforce behavioral reproductive isolation because interspecies courtship is rare even in sexually na{\"ıve animals of most species. We find that the chemoreceptor Gr32a inhibits male D. melanogaster from courting diverse fruit fly species. Gr32a recognizes nonvolatile aversive cues present on these reproductively dead-end targets, and activity of Gr32a neurons is necessary and sufficient to inhibit interspecies courtship. Male-specific Fruitless (Fru(M)), a master regulator of courtship, also inhibits interspecies courtship. Gr32a and Fru(M) are not coexpressed, but Fru(M) neurons contact Gr32a neurons, suggesting that these genes influence a shared neural circuit that inhibits interspecies courtship. Gr32a and Fru(M) also suppress within-species intermale courtship, but we show that distinct mechanisms preclude sexual displays toward conspecific males and other species. Although this chemosensory pathway does not inhibit interspecies mating in D. melanogaster females, similar mechanisms appear to inhibit this behavior in many other male drosophilids.
Alpha/Y-type retinal ganglion cells encode visual information with a receptive field composed of nonlinear subunits. This nonlinear subunit structure enhances sensitivity to patterns composed of high spatial frequencies. The Y-cell’s subunits are the presynaptic bipolar cells, but the mechanism for the nonlinearity remains incompletely understood. We investigated the synaptic basis of the subunit nonlinearity by combining whole-cell recording of mouse Y-type ganglion cells with two-photon fluorescence imaging of a glutamate sensor (iGluSnFR) expressed on their dendrites and throughout the inner plexiform layer. A control experiment designed to assess iGluSnFR’s dynamic range showed that fluorescence responses from Y-cell dendrites increased proportionally with simultaneously recorded excitatory current. Spatial resolution was sufficient to readily resolve independent release at intermingled ON and OFF bipolar terminals. iGluSnFR responses at Y-cell dendrites showed strong surround inhibition, reflecting receptive field properties of presynaptic release sites. Responses to spatial patterns located the origin of the Y-cell nonlinearity to the bipolar cell output, after the stage of spatial integration. The underlying mechanism differed between OFF and ON pathways: OFF synapses showed transient release and strong rectification, whereas ON synapses showed relatively sustained release and weak rectification. At ON synapses, the combination of fast release onset with slower release offset explained the nonlinear response of the postsynaptic ganglion cell. Imaging throughout the inner plexiform layer, we found transient, rectified release at the central-most levels, with increasingly sustained release near the borders. By visualizing glutamate release in real time, iGluSnFR provides a powerful tool for characterizing glutamate synapses in intact neural circuits.
Synaptic loss is the cardinal feature linking neuropathology to cognitive decline in Alzheimer’s disease (AD). However, the mechanism of synaptic damage remains incompletely understood. Here, using FRET-based glutamate sensor imaging, we show that amyloid-β peptide (Aβ) engages α7 nicotinic acetylcholine receptors to induce release of astrocytic glutamate, which in turn activates extrasynaptic NMDA receptors (eNMDARs) on neurons. In hippocampal autapses, this eNMDAR activity is followed by reduction in evoked and miniature excitatory postsynaptic currents (mEPSCs). Decreased mEPSC frequency may reflect early synaptic injury because of concurrent eNMDAR-mediated NO production, tau phosphorylation, and caspase-3 activation, each of which is implicated in spine loss. In hippocampal slices, oligomeric Aβ induces eNMDAR-mediated synaptic depression. In AD-transgenic mice compared with wild type, whole-cell recordings revealed excessive tonic eNMDAR activity accompanied by eNMDAR-sensitive loss of mEPSCs. Importantly, the improved NMDAR antagonist NitroMemantine, which selectively inhibits extrasynaptic over physiological synaptic NMDAR activity, protects synapses from Aβ-induced damage both in vitro and in vivo.
BACKGROUND: Diastolic dysfunction is a poorly understood but clinically pervasive syndrome that is characterized by increased diastolic stiffness. Titin is the main determinant of cellular passive stiffness. However, the physiological role that the tandem immunoglobulin (Ig) segment of titin plays in stiffness generation and whether shortening this segment is sufficient to cause diastolic dysfunction need to be established. METHODS AND RESULTS: We generated a mouse model in which 9 Ig-like domains (Ig3-Ig11) were deleted from the proximal tandem Ig segment of the spring region of titin (IG KO). Exon microarray analysis revealed no adaptations in titin splicing, whereas novel phospho-specific antibodies did not detect changes in titin phosphorylation. Passive myocyte stiffness was increased in the IG KO, and immunoelectron microscopy revealed increased extension of the remaining titin spring segments as the sole likely underlying mechanism. Diastolic stiffness was increased at the tissue and organ levels, with no consistent changes in extracellular matrix composition or extracellular matrix-based passive stiffness, supporting a titin-based mechanism for in vivo diastolic dysfunction. Additionally, IG KO mice have a reduced exercise tolerance, a phenotype often associated with diastolic dysfunction. CONCLUSIONS: Increased titin-based passive stiffness is sufficient to cause diastolic dysfunction with exercise intolerance.
Detection of protein homology via sequence similarity has important applications in biology, from protein structure and function prediction to reconstruction of phylogenies. Although current methods for aligning protein sequences are powerful, challenges remain, including problems with homologous overextension of alignments and with regions under convergent evolution. Here, we test the ability of the profile hidden Markov model method HMMER3 to correctly assign homologous sequences to >13,000 manually curated families from the Pfam database. We identify problem families using protein regions that match two or more Pfam families not currently annotated as related in Pfam. We find that HMMER3 E-value estimates seem to be less accurate for families that feature periodic patterns of compositional bias, such as the ones typically observed in coiled-coils. These results support the continued use of manually curated inclusion thresholds in the Pfam database, especially on the subset of families that have been identified as problematic in experiments such as these. They also highlight the need for developing new methods that can correct for this particular type of compositional bias.
A key step toward understanding a metagenomics data set is the identification of functional sequence elements within it, such as protein coding genes and structural RNAs. Relative to protein coding genes, structural RNAs are more difficult to identify because of their reduced alphabet size, lack of open reading frames, and short length. Infernal is a software package that implements "covariance models" (CMs) for RNA homology search, which harness both sequence and structural conservation when searching for RNA homologs. Thanks to the added statistical signal inherent in the secondary structure conservation of many RNA families, Infernal is more powerful than sequence-only based methods such as BLAST and profile HMMs. Together with the Rfam database of CMs, Infernal is a useful tool for identifying RNAs in metagenomics data sets.
In vertebrates, primary sex determination refers to the decision within a bipotential organ precursor to differentiate as a testis or ovary. Bifurcation of organ fate begins between embryonic day (E) 11.0–E12.0 in mice and likely involves a dynamic transcription network that is poorly understood. To elucidate the first steps of sexual fate specification, we profiled the XX and XY gonad transcriptomes at fine granularity during this period and resolved cascades of gene activation and repression. C57BL/6J (B6) XY gonads showed a consistent 5-hour delay in the activation of most male pathway genes and repression of female pathway genes relative to 129S1/SvImJ, which likely explains the sensitivity of the B6 strain to male-to-female sex reversal. Using this fine time course data, we predicted novel regulatory genes underlying expression QTLs (eQTLs) mapped in a previous study. To test predictions, we developed an in vitro gonad primary cell assay and optimized a lentivirus-based shRNA delivery method to silence candidate genes and quantify effects on putative targets. We provide strong evidence that Lmo4 (Lim-domain only 4) is a novel regulator of sex determination upstream of SF1 (Nr5a1), Sox9, Fgf9, and Col9a3. This approach can be readily applied to identify regulatory interactions in other systems.