Filter
Associated Lab
- Ahrens Lab (1) Apply Ahrens Lab filter
- Baker Lab (2) Apply Baker Lab filter
- Betzig Lab (3) Apply Betzig Lab filter
- Cardona Lab (6) Apply Cardona Lab filter
- Chklovskii Lab (1) Apply Chklovskii Lab filter
- Dickson Lab (2) Apply Dickson Lab filter
- Druckmann Lab (1) Apply Druckmann Lab filter
- Eddy/Rivas Lab (3) Apply Eddy/Rivas Lab filter
- Fetter Lab (2) Apply Fetter Lab filter
- Gonen Lab (7) Apply Gonen Lab filter
- Grigorieff Lab (6) Apply Grigorieff Lab filter
- Harris Lab (1) Apply Harris Lab filter
- Heberlein Lab (5) Apply Heberlein Lab filter
- Hess Lab (2) Apply Hess Lab filter
- Jayaraman Lab (2) Apply Jayaraman Lab filter
- Ji Lab (1) Apply Ji Lab filter
- Kainmueller Lab (2) Apply Kainmueller Lab filter
- Keller Lab (4) Apply Keller Lab filter
- Lavis Lab (1) Apply Lavis Lab filter
- Lee (Albert) Lab (1) Apply Lee (Albert) Lab filter
- Lippincott-Schwartz Lab (12) Apply Lippincott-Schwartz Lab filter
- Looger Lab (7) Apply Looger Lab filter
- Magee Lab (3) Apply Magee Lab filter
- Pastalkova Lab (1) Apply Pastalkova Lab filter
- Reiser Lab (3) Apply Reiser Lab filter
- Riddiford Lab (2) Apply Riddiford Lab filter
- Romani Lab (1) Apply Romani Lab filter
- Rubin Lab (4) Apply Rubin Lab filter
- Saalfeld Lab (5) Apply Saalfeld Lab filter
- Satou Lab (2) Apply Satou Lab filter
- Scheffer Lab (3) Apply Scheffer Lab filter
- Schreiter Lab (2) Apply Schreiter Lab filter
- Shroff Lab (1) Apply Shroff Lab filter
- Simpson Lab (3) Apply Simpson Lab filter
- Singer Lab (1) Apply Singer Lab filter
- Spruston Lab (2) Apply Spruston Lab filter
- Stern Lab (8) Apply Stern Lab filter
- Sternson Lab (1) Apply Sternson Lab filter
- Svoboda Lab (7) Apply Svoboda Lab filter
- Tjian Lab (2) Apply Tjian Lab filter
- Truman Lab (4) Apply Truman Lab filter
- Turaga Lab (2) Apply Turaga Lab filter
- Turner Lab (1) Apply Turner Lab filter
- Zuker Lab (1) Apply Zuker Lab filter
Associated Project Team
Publication Date
- December 2010 (8) Apply December 2010 filter
- November 2010 (9) Apply November 2010 filter
- October 2010 (15) Apply October 2010 filter
- September 2010 (12) Apply September 2010 filter
- August 2010 (12) Apply August 2010 filter
- July 2010 (8) Apply July 2010 filter
- June 2010 (12) Apply June 2010 filter
- May 2010 (6) Apply May 2010 filter
- April 2010 (12) Apply April 2010 filter
- March 2010 (9) Apply March 2010 filter
- February 2010 (16) Apply February 2010 filter
- January 2010 (42) Apply January 2010 filter
- Remove 2010 filter 2010
Type of Publication
161 Publications
Showing 101-110 of 161 resultsPositive-strand RNA [(+)RNA] viruses invariably replicate their RNA genomes on modified intracellular membranes. In infected Drosophila cells, Flock House nodavirus (FHV) RNA replication complexes form on outer mitochondrial membranes inside \~{}50-nm, virus-induced spherular invaginations similar to RNA replication-linked spherules induced by many (+)RNA viruses at various membranes. To better understand replication complex assembly, we studied the mechanisms of FHV spherule formation. FHV has two genomic RNAs; RNA1 encodes multifunctional RNA replication protein A and RNA interference suppressor protein B2, while RNA2 encodes the capsid proteins. Expressing genomic RNA1 without RNA2 induced mitochondrial spherules indistinguishable from those in FHV infection. RNA1 mutation showed that protein B2 was dispensable and that protein A was the only FHV protein required for spherule formation. However, expressing protein A alone only "zippered" together the surfaces of adjacent mitochondria, without inducing spherules. Thus, protein A is necessary but not sufficient for spherule formation. Coexpressing protein A plus a replication-competent FHV RNA template induced RNA replication in trans and membrane spherules. Moreover, spherules were not formed when replicatable FHV RNA templates were expressed with protein A bearing a single, polymerase-inactivating amino acid change or when wild-type protein A was expressed with a nonreplicatable FHV RNA template. Thus, unlike many (+)RNA viruses, the membrane-bounded compartments in which FHV RNA replication occurs are not induced solely by viral protein(s) but require viral RNA synthesis. In addition to replication complex assembly, the results have implications for nodavirus interaction with cell RNA silencing pathways and other aspects of virus control.
The internal workings of the nucleus remain a mystery. A list of component parts exists, and in many cases their functional roles are known for events such as transcription, RNA processing, or nuclear export. Some of these components exhibit structural features in the nucleus, regions of concentration or bodies that have given rise to the concept of functional compartmentalization–that there are underlying organizational principles to be described. In contrast, a picture is emerging in which transcription appears to drive the assembly of the functional components required for gene expression, drawing from pools of excess factors. Unifying this seemingly dual nature requires a more rigorous approach, one in which components are tracked in time and space and correlated with onset of specific nuclear functions. In this chapter, we anticipate tools that will address these questions and provide the missing kinetics of nuclear function. These tools are based on analyzing the fluctuations inherent in the weak signals of endogenous nuclear processes and determining values for them. In this way, it will be possible eventually to provide a computational model describing the functional relationships of essential components.
A striking aspect of cortical neural networks is the divergence of a relatively small number of input channels from the peripheral sensory apparatus into a large number of cortical neurons, an over-complete representation strategy. Cortical neurons are then connected by a sparse network of lateral synapses. Here we propose that such architecture may increase the persistence of the representation of an incoming stimulus, or a percept. We demonstrate that for a family of networks in which the receptive field of each neuron is re-expressed by its outgoing connections, a represented percept can remain constant despite changing activity. We term this choice of connectivity REceptive FIeld REcombination (REFIRE) networks. The sparse REFIRE network may serve as a high-dimensional integrator and a biologically plausible model of the local cortical circuit.
Genes include cis-regulatory regions that contain transcriptional enhancers. Recent reports have shown that developmental genes often possess multiple discrete enhancer modules that drive transcription in similar spatio-temporal patterns: primary enhancers located near the basal promoter and secondary, or ’shadow’, enhancers located at more remote positions. It has been proposed that the seemingly redundant activity of primary and secondary enhancers contributes to phenotypic robustness. We tested this hypothesis by generating a deficiency that removes two newly discovered enhancers of shavenbaby (svb, a transcript of the ovo locus), a gene encoding a transcription factor that directs development of Drosophila larval trichomes. At optimal temperatures for embryonic development, this deficiency causes minor defects in trichome patterning. In embryos that develop at both low and high extreme temperatures, however, absence of these secondary enhancers leads to extensive loss of trichomes. These temperature-dependent defects can be rescued by a transgene carrying a secondary enhancer driving transcription of the svb cDNA. Finally, removal of one copy of wingless, a gene required for normal trichome patterning, causes a similar loss of trichomes only in flies lacking the secondary enhancers. These results support the hypothesis that secondary enhancers contribute to phenotypic robustness in the face of environmental and genetic variability.
In mammals, cerebellar neurons are categorized as glutamatergic or GABAergic, and are derived from progenitors that express the proneural genes atoh1 or ptf1a, respectively. In zebrafish, three atoh1 genes, atoh1a, atoh1b, and atoh1c, are expressed in overlapping but distinct expression domains in the upper rhombic lip (URL): ptf1a is expressed exclusively in the ventricular zone (VZ). Using transgenic lines expressing fluorescent proteins under the control of the regulatory elements of atoh1a and ptf1a, we traced the lineages of the cerebellar neurons. The atoh1+ progenitors gave rise not only to granule cells but also to neurons of the anteroventral rhombencephalon. The ptf1a+ progenitors generated Purkinje cells. The olig2+ eurydendroid cells, which are glutamatergic, were derived mostly from ptf1a+ progenitors in the VZ but some originated from the atoh1+ progenitors in the URL. In the adult cerebellum, atoh1a, atoh1b, and atoh1c are expressed in the molecular layer of the valvula cerebelli and of the medial corpus cerebelli, and ptf1a was detected in the VZ. The proneural gene expression patterns coincided with the sites of proliferating neuronal progenitors in the adult cerebellum. Our data indicate that proneural gene-linked neurogenesis is evolutionarily conserved in the cerebellum among vertebrates, and that the continuously generated neurons help remodel neural circuits in the adult zebrafish cerebellum.
There is considerable interest in the regulation of sensorimotor gating, since deficits in this process could play a critical role in the symptoms of schizophrenia and other psychiatric disorders. Sensorimotor gating is often studied in humans and rodents using the prepulse inhibition of the acoustic startle response (PPI) model, in which an acoustic prepulse suppresses behavioral output to a startle-inducing stimulus. However, the molecular and neural mechanisms underlying PPI are poorly understood. Here, we show that a regulatory pathway involving protein phosphatase 2A (PP2A), glycogen synthase kinase 3 beta (GSK3beta), and their downstream target, the M-type potassium channel, regulates PPI. Mice (Mus musculus) carrying a hypomorphic allele of Ppp2r5delta, encoding a regulatory subunit of PP2A, show attenuated PPI. This PPP2R5delta reduction increases the phosphorylation of GSK3beta at serine 9, which inactivates GSK3beta, indicating that PPP2R5delta positively regulates GSK3beta activity in the brain. Consistently, genetic and pharmacological manipulations that reduce GSK3beta function attenuate PPI. The M-type potassium channel subunit, KCNQ2, is a putative GSK3beta substrate. Genetic reduction of Kcnq2 also reduces PPI, as does systemic inhibition of M-channels with linopirdine. Importantly, both the GSK3 inhibitor 3-(2,4-dichlorophenyl)-4-(1-methyl-1H-indol-3-yl)1H-pyrrole-2,5-dione (SB216763) and linopirdine reduce PPI when directly infused into the medial prefrontal cortex (mPFC). Whole-cell electrophysiological recordings of mPFC neurons show that SB216763 and linopirdine have similar effects on firing, and GSK3 inhibition occludes the effects of M-channel inhibition. These data support a previously uncharacterized mechanism by which PP2A/GSK3beta signaling regulates M-type potassium channel activity in the mPFC to modulate sensorimotor gating.
Recently it was demonstrated that long-lived quantum coherence exists during excitation energy transport in photosynthesis. It is a valid question up to which length, time and mass scales quantum coherence may extend, how one may detect this coherence and what, if any, role it plays in the dynamics of the system. Here we suggest that the selectivity filter of ion channels may exhibit quantum coherence, which might be relevant for the process of ion selectivity and conduction. We show that quantum resonances could provide an alternative approach to ultrafast two-dimensional (2D) spectroscopy to probe these quantum coherences. We demonstrate that the emergence of resonances in the conduction of ion channels that are modulated periodically by time-dependent external electric fields can serve as signatures of quantum coherence in such a system. Assessments of experimental feasibility and specific paths towards the experimental realization of such experiments are presented.
According to spike-timing-dependent plasticity (STDP), the timing of the Na(+) spike relative to the EPSP determines whether LTP or LTD will occur. Here, we review our reservations about STDP. Most investigations of this process have been done under conditions in which the spike is evoked by postsynaptic current injection. Under more realistic conditions, in which the spike is evoked by the EPSP, the results do not generally support STDP. For instance, low-frequency stimulation of a group of synapses can cause LTD, not the LTP predicted by the pre-before-post sequence in STDP; this is true regardless of whether or not the EPSP is large enough to produce a Na(+) spike. With stronger or more frequent stimulation, LTP can be induced by the same pre-before-post timing, but in this case block of Na(+) spikes does not necessarily prevent LTP induction. Thus, Na(+) spikes may facilitate LTP and/or LTD under some conditions, but they are not necessary, a finding consistent with their small size relative to the EPSP in many parts of pyramidal cell dendrites. The nature of the dendritic depolarizing events that control bidirectional plasticity is of central importance to understanding neural function. There are several candidates, including backpropagating action potentials, but also dendritic Ca(2+) spikes, the AMPA receptor-mediated EPSP, and NMDA receptor-mediated EPSPs or spikes. These often appear to be more important than the Na(+) spike in providing the depolarization necessary for plasticity. We thus feel that it is premature to accept STDP-like processes as the major determinant of LTP/LTD.

Novel approaches to bio-imaging and automated computational image processing allow the design of truly quantitative studies in developmental biology. Cell behavior, cell fate decisions, cell interactions during tissue morphogenesis, and gene expression dynamics can be analyzed in vivo for entire complex organisms and throughout embryonic development. We review state-of-the-art technology for live imaging, focusing on fluorescence light microscopy techniques for system-level investigations of animal development and discuss computational approaches to image segmentation, cell tracking, automated data annotation, and biophysical modeling. We argue that the substantial increase in data complexity and size requires sophisticated new strategies to data analysis to exploit the enormous potential of these new resources.
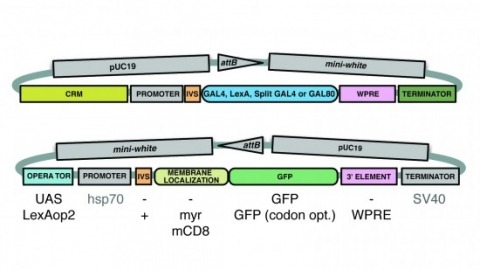
A wide variety of biological experiments rely on the ability to express an exogenous gene in a transgenic animal at a defined level and in a spatially and temporally controlled pattern. We describe major improvements of the methods available for achieving this objective in Drosophila melanogaster. We have systematically varied core promoters, UTRs, operator sequences, and transcriptional activating domains used to direct gene expression with the GAL4, LexA, and Split GAL4 transcription factors and the GAL80 transcriptional repressor. The use of site-specific integration allowed us to make quantitative comparisons between different constructs inserted at the same genomic location. We also characterized a set of PhiC31 integration sites for their ability to support transgene expression of both drivers and responders in the nervous system. The increased strength and reliability of these optimized reagents overcome many of the previous limitations of these methods and will facilitate genetic manipulations of greater complexity and sophistication.