Filter
Associated Lab
- Ahrens Lab (1) Apply Ahrens Lab filter
- Aso Lab (1) Apply Aso Lab filter
- Baker Lab (3) Apply Baker Lab filter
- Betzig Lab (7) Apply Betzig Lab filter
- Bock Lab (2) Apply Bock Lab filter
- Branson Lab (1) Apply Branson Lab filter
- Cardona Lab (1) Apply Cardona Lab filter
- Cui Lab (3) Apply Cui Lab filter
- Dickson Lab (2) Apply Dickson Lab filter
- Druckmann Lab (1) Apply Druckmann Lab filter
- Dudman Lab (1) Apply Dudman Lab filter
- Eddy/Rivas Lab (5) Apply Eddy/Rivas Lab filter
- Fetter Lab (4) Apply Fetter Lab filter
- Fitzgerald Lab (1) Apply Fitzgerald Lab filter
- Gonen Lab (5) Apply Gonen Lab filter
- Grigorieff Lab (6) Apply Grigorieff Lab filter
- Heberlein Lab (12) Apply Heberlein Lab filter
- Hermundstad Lab (1) Apply Hermundstad Lab filter
- Hess Lab (2) Apply Hess Lab filter
- Jayaraman Lab (2) Apply Jayaraman Lab filter
- Ji Lab (2) Apply Ji Lab filter
- Kainmueller Lab (1) Apply Kainmueller Lab filter
- Keller Lab (4) Apply Keller Lab filter
- Lavis Lab (5) Apply Lavis Lab filter
- Lee (Albert) Lab (1) Apply Lee (Albert) Lab filter
- Leonardo Lab (2) Apply Leonardo Lab filter
- Lippincott-Schwartz Lab (18) Apply Lippincott-Schwartz Lab filter
- Liu (Zhe) Lab (1) Apply Liu (Zhe) Lab filter
- Looger Lab (7) Apply Looger Lab filter
- Magee Lab (1) Apply Magee Lab filter
- Menon Lab (3) Apply Menon Lab filter
- Murphy Lab (1) Apply Murphy Lab filter
- Pastalkova Lab (1) Apply Pastalkova Lab filter
- Pavlopoulos Lab (2) Apply Pavlopoulos Lab filter
- Reiser Lab (2) Apply Reiser Lab filter
- Riddiford Lab (1) Apply Riddiford Lab filter
- Romani Lab (1) Apply Romani Lab filter
- Rubin Lab (4) Apply Rubin Lab filter
- Satou Lab (3) Apply Satou Lab filter
- Scheffer Lab (2) Apply Scheffer Lab filter
- Schreiter Lab (3) Apply Schreiter Lab filter
- Sgro Lab (2) Apply Sgro Lab filter
- Simpson Lab (3) Apply Simpson Lab filter
- Singer Lab (10) Apply Singer Lab filter
- Spruston Lab (1) Apply Spruston Lab filter
- Stern Lab (4) Apply Stern Lab filter
- Sternson Lab (6) Apply Sternson Lab filter
- Svoboda Lab (7) Apply Svoboda Lab filter
- Tjian Lab (4) Apply Tjian Lab filter
- Truman Lab (1) Apply Truman Lab filter
- Turaga Lab (1) Apply Turaga Lab filter
- Turner Lab (2) Apply Turner Lab filter
- Zlatic Lab (1) Apply Zlatic Lab filter
- Zuker Lab (3) Apply Zuker Lab filter
Associated Project Team
Publication Date
- December 2011 (22) Apply December 2011 filter
- November 2011 (15) Apply November 2011 filter
- October 2011 (14) Apply October 2011 filter
- September 2011 (17) Apply September 2011 filter
- August 2011 (14) Apply August 2011 filter
- July 2011 (10) Apply July 2011 filter
- June 2011 (17) Apply June 2011 filter
- May 2011 (13) Apply May 2011 filter
- April 2011 (11) Apply April 2011 filter
- March 2011 (14) Apply March 2011 filter
- February 2011 (16) Apply February 2011 filter
- January 2011 (27) Apply January 2011 filter
- Remove 2011 filter 2011
Type of Publication
190 Publications
Showing 171-180 of 190 resultsThe vertebrate hindbrain contains various sensory-motor networks controlling movements of the eyes, jaw, head, and body. Here we show that stripes of neurons with shared neurotransmitter phenotype that extend throughout the hindbrain of young zebrafish reflect a broad underlying structural and functional patterning. The neurotransmitter stripes contain cell types with shared gross morphologies and transcription factor markers. Neurons within a stripe are stacked systematically by extent and location of axonal projections, input resistance, and age, and are recruited along the axis of the stripe during behavior. The implication of this pattern is that the many networks in hindbrain are constructed from a series of neuronal components organized into stripes that are ordered from top to bottom according to a neuron's age, structural and functional properties, and behavioral roles. This simple organization probably forms a foundation for the construction of the networks underlying the many behaviors produced by the hindbrain.
The biogenesis of a localization-competent mRNP begins in the nucleus. It is thought that the coordinated action of nuclear and cytoplasmic components of the localization machinery is required for the efficient export and subsequent subcellular localization of these mRNAs in the cytoplasm. Using quantitative poly(A)(+) and transcript-specific fluorescent in situ hybridization, we analyzed different nonessential nucleoporins and nuclear pore-associated proteins for their potential role in mRNA export and localization. We found that Nup60p, a nuclear pore protein located on the nucleoplasmic side of the nuclear pore complex, was required for the mRNA localization pathway. In a Δnup60 background, localized mRNAs were preferentially retained within the nucleus compared to nonlocalized transcripts. However, the export block was only partial and some transcripts could still reach the cytoplasm. Importantly, downstream processes were also affected. Localization of ASH1 and IST2 mRNAs to the bud was impaired in the Δnup60 background, suggesting that the assembly of a localization competent mRNP ("locasome") was inhibited when NUP60 was deleted. These results demonstrate transcript specificity of a nuclear mRNA retention defect and identify a specific nucleoporin as a functional component of the localization pathway in budding yeast.
Motivation: Digital reconstruction, or tracing, of 3D neuron structures is critical toward reverse engineering the wiring and functions of a brain. However, despite a number of existing studies, this task is still challenging, especially when a 3D microscopic image has low signal-to-noise ratio (SNR) and fragmented neuron segments. Published work can handle these hard situations only by introducing global prior information, such as where a neurite segment starts and terminates. However, manual incorporation of such global information can be very time consuming. Thus, a completely automatic approach for these hard situations is highly desirable. Results: We have developed an automatic graph algorithm, called the all-path pruning (APP), to trace the 3D structure of a neuron. To avoid potential mis-tracing of some parts of a neuron, an APP first produces an initial over-reconstruction, by tracing the optimal geodesic shortest path from the seed location to every possible destination voxel/pixel location in the image. Since the initial reconstruction contains all the possible paths and thus could contain redundant structural components (SC), we simplify the entire reconstruction without compromising its connectedness by pruning the redundant structural elements, using a new maximal-covering minimal-redundant (MCMR) subgraph algorithm. We show that MCMR has a linear computational complexity and will converge. We examined the performance of our method using challenging 3D neuronal image datasets of model organisms (e.g. fruit fly).
Understanding how cells of all types sense external and internal signals and how these signals are processed to yield particular responses is a major goal of biology. Genetically encoded fluorescent proteins (FPs) and fluorescent sensors are playing an important role in achieving this comprehensive knowledge base of cell function. Providing high sensitivity and immense versatility while being minimally perturbing to a biological specimen, the probes can be used in different microscopy techniques to visualize cellular processes on many spatial scales. Three review articles in this volume discuss recent advances in probe design and applications. These developments help expand the range of biochemical processes in living systems suitable for study. They provide researchers with exciting new tools to explore how cellular processes are organized and their activity regulated in vivo.
Optogenetics is routinely used to activate and inactivate genetically defined neuronal populations in vivo. A second optogenetic revolution will occur when spatially distributed and sparse neural assemblies can be precisely manipulated in behaving animals.
The challenge of recovering the topology of massive neuronal circuits can potentially be met by high throughput Electron Microscopy (EM) imagery. Segmenting a 3-dimensional stack of EM images into the individual neurons is difficult, due to the low depth-resolution in existing high-throughput EM technology, such as serial section Transmission EM (ssTEM). In this paper we propose methods for detecting the high resolution locations of membranes from low depth-resolution images. We approach this problem using both a method that learns a discriminative, over-complete dictionary and a kernel SVM. We test this approach on tomographic sections produced in simulations from high resolution Focused Ion Beam (FIB) images and on low depth-resolution images acquired with ssTEM and evaluate our results by comparing it to manual labeling of this data.
Genetically encoded calcium indicators (GECIs), which are based on chimeric fluorescent proteins, can be used to monitor calcium transients in living cells and organisms. Because they are encoded by DNA, GECIs can be delivered to the intact brain noninvasively and targeted to defined populations of neurons and specific subcellular compartments for long-term, repeated measurements in vivo. GECIs have improved iteratively and are becoming useful for imaging neural activity in vivo. Here we summarize extrinsic and intrinsic factors that influence a GECI's performance and provides guidelines for selecting the appropriate GECI for a given application. We also review recent progress in GECI design, optimization, and standardized testing protocols.
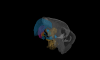
The basal ganglia play a critical role in the regulation of voluntary action in vertebrates. Our understanding of the function of the basal ganglia relies heavily upon anatomical information, but continued progress will require an understanding of the specific functional roles played by diverse cell types and their connectivity. An increasing number of mouse lines allow extensive identification, characterization, and manipulation of specified cell types in the basal ganglia. Despite the promise of genetically modified mice for elucidating the functional roles of diverse cell types, there is relatively little anatomical data obtained directly in the mouse. Here we have characterized the retrograde labeling obtained from a series of tracer injections throughout the dorsal striatum of adult mice. We found systematic variations in input along both the medial-lateral and anterior-posterior neuraxes in close agreement with canonical features of basal ganglia anatomy in the rat. In addition to the canonical features we have provided experimental support for the importance of non-canonical inputs to the striatum from the raphe nuclei and the amygdala. To look for organization at a finer scale we have analyzed the correlation structure of labeling intensity across our entire dataset. Using this analysis we found substantial local heterogeneity within the large-scale order. From this analysis we conclude that individual striatal sites receive varied combinations of cortical and thalamic input from multiple functional areas, consistent with some earlier studies in the rat that have suggested the presence of a combinatorial map.
An agglomerative clustering algorithm merges the most similar pair of clusters at every iteration. The function that evaluates similarity is traditionally handdesigned, but there has been recent interest in supervised or semisupervised settings in which ground-truth clustered data is available for training. Here we show how to train a similarity function by regarding it as the action-value function of a reinforcement learning problem. We apply this general method to segment images by clustering superpixels, an application that we call Learning to Agglomerate Superpixel Hierarchies (LASH). When applied to a challenging dataset of brain images from serial electron microscopy, LASH dramatically improved segmentation accuracy when clustering supervoxels generated by state of the boundary detection algorithms. The naive strategy of directly training only supervoxel similarities and applying single linkage clustering produced less improvement.