Filter
Associated Lab
- Ahrens Lab (1) Apply Ahrens Lab filter
- Aso Lab (1) Apply Aso Lab filter
- Baker Lab (3) Apply Baker Lab filter
- Betzig Lab (7) Apply Betzig Lab filter
- Bock Lab (2) Apply Bock Lab filter
- Branson Lab (1) Apply Branson Lab filter
- Cardona Lab (1) Apply Cardona Lab filter
- Cui Lab (3) Apply Cui Lab filter
- Dickson Lab (2) Apply Dickson Lab filter
- Druckmann Lab (1) Apply Druckmann Lab filter
- Dudman Lab (1) Apply Dudman Lab filter
- Eddy/Rivas Lab (5) Apply Eddy/Rivas Lab filter
- Fetter Lab (4) Apply Fetter Lab filter
- Fitzgerald Lab (1) Apply Fitzgerald Lab filter
- Gonen Lab (5) Apply Gonen Lab filter
- Grigorieff Lab (6) Apply Grigorieff Lab filter
- Heberlein Lab (12) Apply Heberlein Lab filter
- Hermundstad Lab (1) Apply Hermundstad Lab filter
- Hess Lab (2) Apply Hess Lab filter
- Jayaraman Lab (2) Apply Jayaraman Lab filter
- Ji Lab (2) Apply Ji Lab filter
- Kainmueller Lab (1) Apply Kainmueller Lab filter
- Keller Lab (4) Apply Keller Lab filter
- Lavis Lab (5) Apply Lavis Lab filter
- Lee (Albert) Lab (1) Apply Lee (Albert) Lab filter
- Leonardo Lab (2) Apply Leonardo Lab filter
- Lippincott-Schwartz Lab (18) Apply Lippincott-Schwartz Lab filter
- Liu (Zhe) Lab (1) Apply Liu (Zhe) Lab filter
- Looger Lab (7) Apply Looger Lab filter
- Magee Lab (1) Apply Magee Lab filter
- Menon Lab (3) Apply Menon Lab filter
- Murphy Lab (1) Apply Murphy Lab filter
- Pastalkova Lab (1) Apply Pastalkova Lab filter
- Pavlopoulos Lab (2) Apply Pavlopoulos Lab filter
- Reiser Lab (2) Apply Reiser Lab filter
- Riddiford Lab (1) Apply Riddiford Lab filter
- Romani Lab (1) Apply Romani Lab filter
- Rubin Lab (4) Apply Rubin Lab filter
- Satou Lab (3) Apply Satou Lab filter
- Scheffer Lab (2) Apply Scheffer Lab filter
- Schreiter Lab (3) Apply Schreiter Lab filter
- Sgro Lab (2) Apply Sgro Lab filter
- Simpson Lab (3) Apply Simpson Lab filter
- Singer Lab (10) Apply Singer Lab filter
- Spruston Lab (1) Apply Spruston Lab filter
- Stern Lab (4) Apply Stern Lab filter
- Sternson Lab (6) Apply Sternson Lab filter
- Svoboda Lab (7) Apply Svoboda Lab filter
- Tjian Lab (4) Apply Tjian Lab filter
- Truman Lab (1) Apply Truman Lab filter
- Turaga Lab (1) Apply Turaga Lab filter
- Turner Lab (2) Apply Turner Lab filter
- Zlatic Lab (1) Apply Zlatic Lab filter
- Zuker Lab (3) Apply Zuker Lab filter
Associated Project Team
Publication Date
- December 2011 (22) Apply December 2011 filter
- November 2011 (15) Apply November 2011 filter
- October 2011 (14) Apply October 2011 filter
- September 2011 (17) Apply September 2011 filter
- August 2011 (14) Apply August 2011 filter
- July 2011 (10) Apply July 2011 filter
- June 2011 (17) Apply June 2011 filter
- May 2011 (13) Apply May 2011 filter
- April 2011 (11) Apply April 2011 filter
- March 2011 (14) Apply March 2011 filter
- February 2011 (16) Apply February 2011 filter
- January 2011 (27) Apply January 2011 filter
- Remove 2011 filter 2011
Type of Publication
190 Publications
Showing 51-60 of 190 resultsThe task of the visual system is to translate light into neuronal encoded information. This translation of photons into neuronal signals is achieved by photoreceptor neurons (PRs), specialized sensory neurons, located in the eye. Upon perception of light the PRs will send a signal to target neurons, which represent a first station of visual processing. Increasing complexity of visual processing stems from the number of distinct PR subtypes and their various types of target neurons that are contacted. The visual system of the fruit fly larva represents a simple visual system (larval optic neuropil, LON) that consists of 12 PRs falling into two classes: blue-senstive PRs expressing Rhodopsin 5 (Rh5) and green-sensitive PRs expressing Rhodopsin 6 (Rh6). These afferents contact a small number of target neurons, including optic lobe pioneers (OLPs) and lateral clock neurons (LNs). We combine the use of genetic markers to label both PR subtypes and the distinct, identifiable sets of target neurons with a serial EM reconstruction to generate a high-resolution map of the larval optic neuropil. We find that the larval optic neuropil shows a clear bipartite organization consisting of one domain innervated by PRs and one devoid of PR axons. The topology of PR projections, in particular the relationship between Rh5 and Rh6 afferents, is maintained from the nerve entering the brain to the axon terminals. The target neurons can be subdivided according to neurotransmitter or neuropeptide they use as well as the location within the brain. We further track the larval optic neuropil through development from first larval instar to its location in the adult brain as the accessory medulla.
Disrupted-in-Schizophrenia-1 (DISC1) is a genetic susceptibility locus for major mental illness, including schizophrenia and depression. The Disc1 protein was recently shown to interact with the Wnt signaling protein, DIX domain containing 1 (Dixdc1). Both proteins participate in neural progenitor proliferation dependent on Wnt signaling, and in neural migration independently of Wnt signaling. Interestingly, their effect on neural progenitor proliferation is additive. By analogy to Disc1, mutations in Dixdc1 may lead to abnormal behavior in mice, and to schizophrenia or depression in humans. To explore this hypothesis further, we generated mice mutant at the Dixdc1 locus and analyzed their behavior. Dixdc1(-/-) mice had normal prepulse inhibition, but displayed decreased spontaneous locomotor activity, abnormal behavior in the elevated plus maze and deficits in startle reactivity. Our results suggest that Dixdc1(-/-) mice will be a useful tool to elucidate molecular pathophysiology involving Disc1 in major mental illnesses.
The molecular structure of amyloid fibrils and the mechanism of their formation are of substantial medical and biological importance, but present an ongoing experimental and computational challenge. An early high-resolution view of amyloid-like structure was obtained on amyloid-like crystals of a small fragment of the yeast prion protein Sup35p: the peptide GNNQQNY. As GNNQQNY also forms amyloid-like fibrils under similar conditions, it has been theorized that the crystal’s structural features are shared by the fibrils. Here we apply magic-angle-spinning (MAS) NMR to examine the structure and dynamics of these fibrils. Previously multiple NMR signals were observed for such samples, seemingly consistent with the presence of polymorphic fibrils. Here we demonstrate that peptides with these three distinct conformations instead assemble together into composite protofilaments. Electron microscopy (EM) of the ribbon-like fibrils indicates that these protofilaments combine in differing ways to form striations of variable widths, presenting another level of structural complexity. Structural and dynamic NMR data reveal the presence of highly restricted side-chain conformations involved in interfaces between differently structured peptides, likely comprising interdigitated steric zippers. We outline molecular interfaces that are consistent with the observed EM and NMR data. The rigid and uniform structure of the GNNQQNY crystals is found to contrast distinctly with the more complex structural and dynamic nature of these "composite" amyloid fibrils. These results provide insight into the fibril-crystal distinction and also indicate a necessary caution with respect to the extrapolation of crystal structures to the study of fibril structure and formation.
Photoactivated localization microscopy (PALM) is a powerful approach for investigating protein organization, yet tools for quantitative, spatial analysis of PALM datasets are largely missing. Combining pair-correlation analysis with PALM (PC-PALM), we provide a method to analyze complex patterns of protein organization across the plasma membrane without determination of absolute protein numbers. The approach uses an algorithm to distinguish a single protein with multiple appearances from clusters of proteins. This enables quantification of different parameters of spatial organization, including the presence of protein clusters, their size, density and abundance in the plasma membrane. Using this method, we demonstrate distinct nanoscale organization of plasma-membrane proteins with different membrane anchoring and lipid partitioning characteristics in COS-7 cells, and show dramatic changes in glycosylphosphatidylinositol (GPI)-anchored protein arrangement under varying perturbations. PC-PALM is thus an effective tool with broad applicability for analysis of protein heterogeneity and function, adaptable to other single-molecule strategies.
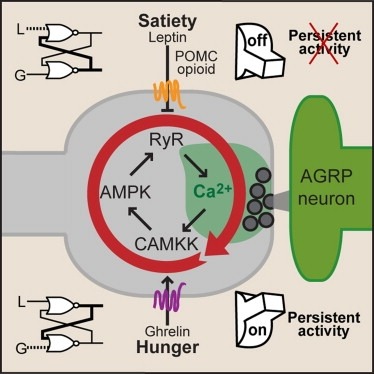
Synaptic plasticity in response to changes in physiologic state is coordinated by hormonal signals across multiple neuronal cell types. Here, we combine cell-type-specific electrophysiological, pharmacological, and optogenetic techniques to dissect neural circuits and molecular pathways controlling synaptic plasticity onto AGRP neurons, a population that regulates feeding. We find that food deprivation elevates excitatory synaptic input, which is mediated by a presynaptic positive feedback loop involving AMP-activated protein kinase. Potentiation of glutamate release was triggered by the orexigenic hormone ghrelin and exhibited hysteresis, persisting for hours after ghrelin removal. Persistent activity was reversed by the anorexigenic hormone leptin, and optogenetic photostimulation demonstrated involvement of opioid release from POMC neurons. Based on these experiments, we propose a memory storage device for physiological state constructed from bistable synapses that are flipped between two sustained activity states by transient exposure to hormones signaling energy levels.
A specialist neuron uses an intriguing process to help control the body's response to hunger. A lipid pathway involving the breakdown of cellular components regulates the expression of a neuropeptide that affects feeding and body weight.
The taste system is one of our fundamental senses, responsible for detecting and responding to sweet, bitter, umami, salty, and sour stimuli. In the tongue, the five basic tastes are mediated by separate classes of taste receptor cells each finely tuned to a single taste quality. We explored the logic of taste coding in the brain by examining how sweet, bitter, umami, and salty qualities are represented in the primary taste cortex of mice. We used in vivo two-photon calcium imaging to demonstrate topographic segregation in the functional architecture of the gustatory cortex. Each taste quality is represented in its own separate cortical field, revealing the existence of a gustotopic map in the brain. These results expose the basic logic for the central representation of taste.
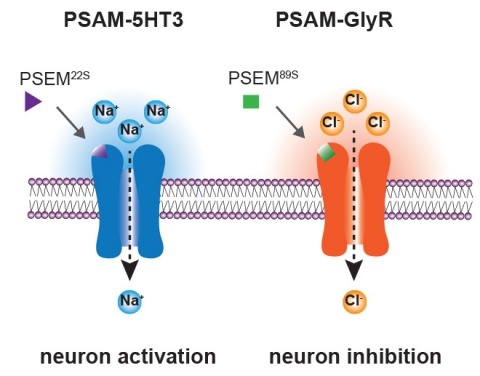
Ionic flux mediates essential physiological and behavioral functions in defined cell populations. Cell type-specific activators of diverse ionic conductances are needed for probing these effects. We combined chemistry and protein engineering to enable the systematic creation of a toolbox of ligand-gated ion channels (LGICs) with orthogonal pharmacologic selectivity and divergent functional properties. The LGICs and their small-molecule effectors were able to activate a range of ionic conductances in genetically specified cell types. LGICs constructed for neuronal perturbation could be used to selectively manipulate neuron activity in mammalian brains in vivo. The diversity of ion channel tools accessible from this approach will be useful for examining the relationship between neuronal activity and animal behavior, as well as for cell biological and physiological applications requiring chemical control of ion conductance.
Deciphering the molecular basis of pluripotency is fundamental to our understanding of development and embryonic stem cell function. Here, we report that TAF3, a TBP-associated core promoter factor, is highly enriched in ES cells. In this context, TAF3 is required for endoderm lineage differentiation and prevents premature specification of neuroectoderm and mesoderm. In addition to its role in the core promoter recognition complex TFIID, genome-wide binding studies reveal that TAF3 localizes to a subset of chromosomal regions bound by CTCF/cohesin that are selectively associated with genes upregulated by TAF3. Notably, CTCF directly recruits TAF3 to promoter distal sites and TAF3-dependent DNA looping is observed between the promoter distal sites and core promoters occupied by TAF3/CTCF/cohesin. Together, our findings support a new role of TAF3 in mediating long-range chromatin regulatory interactions that safeguard the finely-balanced transcriptional programs underlying pluripotency.
Establishing visual correspondences is a critical step in many computer vision tasks involving multiple views of a scene. In a dynamic environment and when cameras are mobile, visual correspondences need to be updated on a recurring basis. At the same time, the use of wireless links between camera motes imposes tight rate constraints. This combination of issues motivates us to consider the problem of establishing visual correspondences in a distributed fashion between cameras operating under rate constraints. We propose a solution based on constructing distance preserving hashes using binarized random projections. By exploiting the fact that descriptors of regions in correspondence are highly correlated, we propose a novel use of distributed source coding via linear codes on the binary hashes to more efficiently exchange feature descriptors for establishing correspondences across multiple camera views. A systematic approach is used to evaluate rate vs visual correspondences retrieval performance; under a stringent matching criterion, our proposed methods demonstrate superior performance to a baseline scheme employing transform coding of descriptors.