Filter
Associated Lab
- Aguilera Castrejon Lab (15) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (11) Apply Ahrens Lab filter
- Baker Lab (19) Apply Baker Lab filter
- Betzig Lab (12) Apply Betzig Lab filter
- Beyene Lab (5) Apply Beyene Lab filter
- Bock Lab (3) Apply Bock Lab filter
- Branson Lab (3) Apply Branson Lab filter
- Card Lab (6) Apply Card Lab filter
- Cardona Lab (19) Apply Cardona Lab filter
- Chklovskii Lab (3) Apply Chklovskii Lab filter
- Clapham Lab (1) Apply Clapham Lab filter
- Darshan Lab (4) Apply Darshan Lab filter
- Dennis Lab (1) Apply Dennis Lab filter
- Dickson Lab (14) Apply Dickson Lab filter
- Druckmann Lab (4) Apply Druckmann Lab filter
- Dudman Lab (12) Apply Dudman Lab filter
- Egnor Lab (7) Apply Egnor Lab filter
- Espinosa Medina Lab (4) Apply Espinosa Medina Lab filter
- Fetter Lab (10) Apply Fetter Lab filter
- Fitzgerald Lab (13) Apply Fitzgerald Lab filter
- Gonen Lab (32) Apply Gonen Lab filter
- Grigorieff Lab (28) Apply Grigorieff Lab filter
- Harris Lab (10) Apply Harris Lab filter
- Heberlein Lab (81) Apply Heberlein Lab filter
- Hermundstad Lab (4) Apply Hermundstad Lab filter
- Hess Lab (3) Apply Hess Lab filter
- Jayaraman Lab (4) Apply Jayaraman Lab filter
- Johnson Lab (5) Apply Johnson Lab filter
- Kainmueller Lab (19) Apply Kainmueller Lab filter
- Karpova Lab (1) Apply Karpova Lab filter
- Keleman Lab (5) Apply Keleman Lab filter
- Keller Lab (15) Apply Keller Lab filter
- Koay Lab (16) Apply Koay Lab filter
- Lavis Lab (12) Apply Lavis Lab filter
- Lee (Albert) Lab (5) Apply Lee (Albert) Lab filter
- Leonardo Lab (4) Apply Leonardo Lab filter
- Li Lab (24) Apply Li Lab filter
- Lippincott-Schwartz Lab (72) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (5) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (5) Apply Liu (Zhe) Lab filter
- Looger Lab (1) Apply Looger Lab filter
- Magee Lab (18) Apply Magee Lab filter
- Menon Lab (6) Apply Menon Lab filter
- Murphy Lab (7) Apply Murphy Lab filter
- O'Shea Lab (1) Apply O'Shea Lab filter
- Otopalik Lab (12) Apply Otopalik Lab filter
- Pachitariu Lab (12) Apply Pachitariu Lab filter
- Pastalkova Lab (13) Apply Pastalkova Lab filter
- Pavlopoulos Lab (12) Apply Pavlopoulos Lab filter
- Pedram Lab (11) Apply Pedram Lab filter
- Reiser Lab (6) Apply Reiser Lab filter
- Riddiford Lab (24) Apply Riddiford Lab filter
- Romani Lab (12) Apply Romani Lab filter
- Rubin Lab (38) Apply Rubin Lab filter
- Saalfeld Lab (17) Apply Saalfeld Lab filter
- Satou Lab (15) Apply Satou Lab filter
- Schreiter Lab (17) Apply Schreiter Lab filter
- Sgro Lab (20) Apply Sgro Lab filter
- Simpson Lab (5) Apply Simpson Lab filter
- Singer Lab (43) Apply Singer Lab filter
- Spruston Lab (36) Apply Spruston Lab filter
- Stern Lab (83) Apply Stern Lab filter
- Sternson Lab (7) Apply Sternson Lab filter
- Stringer Lab (3) Apply Stringer Lab filter
- Svoboda Lab (4) Apply Svoboda Lab filter
- Tebo Lab (24) Apply Tebo Lab filter
- Tillberg Lab (3) Apply Tillberg Lab filter
- Tjian Lab (47) Apply Tjian Lab filter
- Truman Lab (30) Apply Truman Lab filter
- Turaga Lab (12) Apply Turaga Lab filter
- Turner Lab (11) Apply Turner Lab filter
- Wang (Shaohe) Lab (19) Apply Wang (Shaohe) Lab filter
- Wu Lab (1) Apply Wu Lab filter
- Zlatic Lab (2) Apply Zlatic Lab filter
- Zuker Lab (20) Apply Zuker Lab filter
Associated Project Team
Publication Date
- 2024 (1) Apply 2024 filter
- 2023 (1) Apply 2023 filter
- 2022 (26) Apply 2022 filter
- 2021 (19) Apply 2021 filter
- 2020 (19) Apply 2020 filter
- 2019 (25) Apply 2019 filter
- 2018 (26) Apply 2018 filter
- 2017 (31) Apply 2017 filter
- 2016 (18) Apply 2016 filter
- 2015 (57) Apply 2015 filter
- 2014 (46) Apply 2014 filter
- 2013 (58) Apply 2013 filter
- 2012 (78) Apply 2012 filter
- 2011 (92) Apply 2011 filter
- 2010 (100) Apply 2010 filter
- 2009 (102) Apply 2009 filter
- 2008 (100) Apply 2008 filter
- 2007 (85) Apply 2007 filter
- 2006 (89) Apply 2006 filter
- 2005 (67) Apply 2005 filter
- 2004 (57) Apply 2004 filter
- 2003 (58) Apply 2003 filter
- 2002 (39) Apply 2002 filter
- 2001 (28) Apply 2001 filter
- 2000 (29) Apply 2000 filter
- 1999 (14) Apply 1999 filter
- 1998 (18) Apply 1998 filter
- 1997 (16) Apply 1997 filter
- 1996 (10) Apply 1996 filter
- 1995 (18) Apply 1995 filter
- 1994 (12) Apply 1994 filter
- 1993 (10) Apply 1993 filter
- 1992 (6) Apply 1992 filter
- 1991 (11) Apply 1991 filter
- 1990 (11) Apply 1990 filter
- 1989 (6) Apply 1989 filter
- 1988 (1) Apply 1988 filter
- 1987 (7) Apply 1987 filter
- 1986 (4) Apply 1986 filter
- 1985 (5) Apply 1985 filter
- 1984 (2) Apply 1984 filter
- 1983 (2) Apply 1983 filter
- 1982 (3) Apply 1982 filter
- 1981 (3) Apply 1981 filter
- 1980 (1) Apply 1980 filter
- 1979 (1) Apply 1979 filter
- 1976 (2) Apply 1976 filter
- 1973 (1) Apply 1973 filter
- 1970 (1) Apply 1970 filter
- 1967 (1) Apply 1967 filter
Type of Publication
- Remove Non-Janelia filter Non-Janelia
1417 Publications
Showing 1351-1360 of 1417 resultsThe near-field optical interaction between a sharp probe and a sample of interest can be exploited to image, spectroscopically probe, or modify surfaces at a resolution (down to approximately 12 nm) inaccessible by traditional far-field techniques. Many of the attractive features of conventional optics are retained, including noninvasiveness, reliability, and low cost. In addition, most optical contrast mechanisms can be extended to the near-field regime, resulting in a technique of considerable versatility. This versatility is demonstrated by several examples, such as the imaging of nanometric-scale features in mammalian tissue sections and the creation of ultrasmall, magneto-optic domains having implications for highdensity data storage. Although the technique may find uses in many diverse fields, two of the most exciting possibilities are localized optical spectroscopy of semiconductors and the fluorescence imaging of living cells.
Commentary: An overview of our work in near-field optics at the time, after our invention of the adiabatically tapered fiber probe and shear force feedback (see below) led to the first practical near-field scanning optical microscope. In this work, superresolution imaging via absorption, reflectivity, fluorescence, spectroscopy, polarization, and refractive index contrast were all demonstrated. Unlike all far-field superresolution fluorescence methods that were to appear a decade later, near-field microscopy remains the only superresolution technique capable of taking advantage of the full panoply of optical contrast mechanisms.

The argos gene encodes a protein that is required for viability and that regulates the determination of cells in the Drosophila eye. A developmental analysis of argos mutant eyes indicates that the mystery cells, which are usually nonneuronal, are transformed into extra photoreceptors, and that supernumerary cone cells and pigment cells are also recruited. Clonal analysis indicates that argos acts nonautonomously and can diffuse over the range of several cell diameters. Conceptual translation of the argos gene suggests that it encodes a secreted protein.
A distance regulation method has been developed to enhance the reliability, versatility, and ease of use of near-field scanning optical microscopy (NSOM). The method relies on the detection of shear forces between the end of a near-field probe and the sample of interest. The system can be used solely for distance regulation in NSOM, for simultaneous shear force and near-field imaging, or for shear force microscopy alone. In the latter case, uncoated optical fiber probes are found to yield images with consistently high resolution.
Commentary: To exploit the evanescent field that is the source of high resolution in near-field microscopy, the probe must be exceptionally close to the sample: 10 nm away for 30-50 nm resolution. Here we introduced a distance regulation mechanism based on transverse shear forces between the end of a dithered near-field probe and the sample, which permitted even samples of modest topography to be imaged. Simple, reliable, noninvasive, and applicable to a wide range of samples from whole fixed cells to semiconductor devices, shear force microscopy was a key enabling technology for near-field optics, and soon widely implemented.
1. Perforated patch-clamp recordings were made from the three major classes of hippocampal neurons in conventional in vitro slices prepared from adult guinea pigs. This technique provided experimental estimates of passive membrane properties (input resistance, RN, and membrane time constant, tau m) determined in the absence of the leak conductance associated with microelectrode impalement or the washout of cytoplasmic constituents associated with conventional whole-cell recordings. 2. To facilitate comparison of our data with previous results and to determine the passive membrane properties under conditions as physiological as possible, recordings were made at the resting potential, in physiological saline, and without any added blockers of voltage-dependent conductances. 3. Membrane-potential responses to current steps were analyzed, and four criteria were used to identify voltage responses that were the least affected by activation of voltage-dependent conductances. tau m was estimated from the slowest component (tau 0) of multiexponential fits of responses deemed passive by these criteria. RN was estimated from the slope of the linear region in the hyperpolarizing direction of the voltage-current relation. 4. It was not possible to measure purely passive membrane properties that were completely independent of membrane potential in any of the three classes of hippocampal neurons. Changing the membrane potential by constant current injection resulted in changes in RN and tau 0; subthreshold depolarization produced an increase, and hyperpolarization a decrease, in both RN and tau 0 for all three classes of hippocampal neurons. 5. Each of the three classes of hippocampal neurons also displayed a depolarizing "sag" during larger hyperpolarizing voltage transients. To evaluate the effect of the conductances underlying this sag on passive membrane properties, 2-5 mM Cs+ was added to the physiological saline. Extracellular Cs+ effectively blocked the sag in all three classes of hippocampal neurons, but the effect of Cs+ on RN, tau 0, and the voltage dependence of these parameters was unique for each class of neurons. 6. CA1 pyramidal neurons had an RN of 104 +/- 10 (SE) M omega and tau 0 of 28 +/- 2 ms at a resting potential of -64 +/- 2 mV (n = 12). RN and tau 0 were larger at more depolarized potentials in these neurons, but the addition of Cs+ to the physiological saline reversed this voltage dependence. 7. CA3 pyramidal neurons had an RN of 135 +/- 8 M omega and tau 0 of 66 +/- 4 ms at a resting potential of -64 +/- 1 mV (n = 14).(ABSTRACT TRUNCATED AT 400 WORDS)
Near-field scanning optical microscopy (NSOM) has been used to image and record domains in thin-film magneto-optic (MO) materials. In the imaging mode, resolution of 30-50 nm has been consistently obtained, whereas in the recording mode, domains down to -60 nm have been written reproducibly. Data densities of -45 Gbits/in.’ have been achieved, well in excess of current magnetic or MO technologies. A brief analysis of speed and other issues indicates that the technique may represent a viable alternative to density data storage needs.
Commentary: The first demonstration of optical recording and playback beyond the diffraction limit, using magneto-optic multilayer films and polarization contrast near-field microscopy. Bits as small as 60 nm were recorded – beyond estimates at the time of the superparamagnetic limit to bit stability. Bit densities of 45 Gbits/in2 were also achieved, well in excess of optical or magnetic recording technologies of the era. In the years following this work, massive resources were spent on the commercialization of near-field data storage, largely for naught.
Examined kin discrimination and colony defense in soldier-producing aphids from the surface of 3 galls. Soldiers always attacked conspecific nonsoldiers, regardless of origin, and never attacked conspecific soldiers. Soldier attacks of nonsoldiers may exclude unrelated nonsoldier aphids from the gall where they would propagate and compete with resident aphids. (PsycINFO Database Record (c) 2012 APA, all rights reserved)

We have conducted a genetic screen for mutations that decrease the effectiveness of signaling by a protein tyrosine kinase, the product of the Drosophila melanogaster sevenless gene. These mutations define seven genes whose wild-type products may be required for signaling by sevenless. Four of the seven genes also appear to be essential for signaling by a second protein tyrosine kinase, the product of the Ellipse gene. The putative products of two of these seven genes have been identified. One encodes a ras protein. The other locus encodes a protein that is homologous to the S. cerevisiae CDC25 protein, an activator of guanine nucleotide exchange by ras proteins. These results suggest that the stimulation of ras protein activity is a key element in the signaling by sevenless and Ellipse and that this stimulation may be achieved by activating the exchange of GTP for bound GDP by the ras protein.
During a four month study of male territoriality males of the euglossine bee Eulaema meriana exhibited the two alternative behavior patterns of territoriality and transiency. Territorial males patrolled an area adjacent to a tree upon which they perched. Territorial males utilized the same territory for up to 49 days, though often not on consecutive days, and appeared to non-violently relinquish territories to new males. Transients did not defend territories but flew from one territory to another and flew with the territorial male around the territory, rarely bumping, and never grappling. Transient males left the territory soon after the territorial male flew back and forth in front of the perch tree in a zig-zag flight. The alternative behaviors were correlated with wing wear such that males with little wing wear defended territories and males with considerable wing wear pursued a transient strategy. Behavior patterns were not correlated with head width. Comparison of territory trees with the territory trees of a closely related species indicate that each species utilized trees of a certain diameter class for perching. In addition, analysis of hemispherical canopy photographs indicates that males appeared to prefer territories that received a maximum of diffuse sunlight but a minimum of direct sunlight. Both territorial and transient males consistently returned to specific territories over their lifetime but appeared to travel long distances to forage for fragrances. Territorial and transient males visited fragrance baits with equal frequency suggesting that non-territorial, as well as territorial, males required fragrances.
The maleless (mle) gene is one of four known regulatory loci required for increased transcription (dosage compensation) of X-linked genes in D. melanogaster males. A predicted mle protein (MLE) contains seven short segments that define a superfamily of known and putative RNA and DNA helicases. MLE, while present in the nuclei of both male and female cells, differs in its association with polytene X chromosomes in the two sexes. MLE is associated with hundreds of discrete sites along the length of the X chromosome in males and not in females. The predominant localization of MLE to the X chromosome in males makes it a strong candidate to be a direct regulator of dosage compensation.
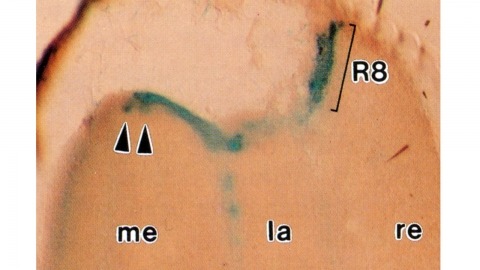
Histological staining of wild-type and sevenless transgenic Drosophila melanogaster bearing Rh3-lacZ fusion genes permits the selective visualization of polarization-sensitive R7 and R8 photoreceptor cells located along the dorsal anterior eye margin. Diffusion of beta-galactosidase throughout these cells reveals that they project long axons to the two most peripheral synaptic target rows of the dorsal posterior medulla, defining a specialized marginal zone of this optic lobe. Comparison of the staining patterns of marginal and nonmarginal Rh3-lacZ-expressing photoreceptor cells in the same histological preparations suggest that the marginal cells possess morphologically specialized axons and synaptic terminals. These findings are discussed with reference to the neuroanatomy of the corresponding dorsal marginal eye and optic lobe regions of the larger dipterans Musca and Calliphora, and in relation to the ability of Drosophila to orient to polarized light.