Filter
Associated Lab
- Ahrens Lab (3) Apply Ahrens Lab filter
- Betzig Lab (8) Apply Betzig Lab filter
- Card Lab (2) Apply Card Lab filter
- Cardona Lab (1) Apply Cardona Lab filter
- Druckmann Lab (1) Apply Druckmann Lab filter
- Eddy/Rivas Lab (2) Apply Eddy/Rivas Lab filter
- Fetter Lab (4) Apply Fetter Lab filter
- Fitzgerald Lab (1) Apply Fitzgerald Lab filter
- Gonen Lab (6) Apply Gonen Lab filter
- Harris Lab (1) Apply Harris Lab filter
- Heberlein Lab (3) Apply Heberlein Lab filter
- Hess Lab (2) Apply Hess Lab filter
- Ji Lab (4) Apply Ji Lab filter
- Kainmueller Lab (1) Apply Kainmueller Lab filter
- Keller Lab (3) Apply Keller Lab filter
- Lavis Lab (4) Apply Lavis Lab filter
- Looger Lab (3) Apply Looger Lab filter
- Magee Lab (4) Apply Magee Lab filter
- Murphy Lab (1) Apply Murphy Lab filter
- Pastalkova Lab (3) Apply Pastalkova Lab filter
- Reiser Lab (1) Apply Reiser Lab filter
- Riddiford Lab (5) Apply Riddiford Lab filter
- Romani Lab (2) Apply Romani Lab filter
- Rubin Lab (4) Apply Rubin Lab filter
- Saalfeld Lab (2) Apply Saalfeld Lab filter
- Satou Lab (1) Apply Satou Lab filter
- Schreiter Lab (3) Apply Schreiter Lab filter
- Sgro Lab (1) Apply Sgro Lab filter
- Shroff Lab (6) Apply Shroff Lab filter
- Spruston Lab (3) Apply Spruston Lab filter
- Stern Lab (4) Apply Stern Lab filter
- Sternson Lab (1) Apply Sternson Lab filter
- Svoboda Lab (6) Apply Svoboda Lab filter
- Tjian Lab (7) Apply Tjian Lab filter
- Truman Lab (2) Apply Truman Lab filter
- Turner Lab (1) Apply Turner Lab filter
- Wu Lab (1) Apply Wu Lab filter
- Zuker Lab (1) Apply Zuker Lab filter
Publication Date
- Remove 2007-12-31 19:00 – 2008-12-31 19:00 filter 2007-12-31 19:00 – 2008-12-31 19:00
- December 2008 (11) Apply December 2008 filter
- November 2008 (8) Apply November 2008 filter
- October 2008 (7) Apply October 2008 filter
- September 2008 (12) Apply September 2008 filter
- August 2008 (10) Apply August 2008 filter
- July 2008 (14) Apply July 2008 filter
- June 2008 (13) Apply June 2008 filter
- May 2008 (8) Apply May 2008 filter
- April 2008 (7) Apply April 2008 filter
- March 2008 (16) Apply March 2008 filter
- February 2008 (13) Apply February 2008 filter
- January 2008 (21) Apply January 2008 filter
Type of Publication
140 Publications
Showing 71-80 of 140 resultsAquaporins (AQPs) are a family of ubiquitous membrane channels that conduct water and solutes across membranes. This review focuses on AQP0 and AQP4, which in addition to forming water channels also appear to play a role in cell adhesion. We discuss the recently determined structures of the membrane junctions mediated by these two AQPs, the mechanisms that regulate junction formation, and evidence that supports a role for AQP0 and AQP4 in cell adhesion.
Juvenile hormone (JH) is a key hormone in regulation of the insect’s life history, both in maintaining the larval state during molts and in directing reproductive maturation. This short review highlights the recent papers of the past year that lend new insight into the role of this hormone in the larva and the mechanisms whereby it achieves this role.
Juvenile hormone (JH) given at pupariation inhibits bristle formation and causes pupal cuticle formation in the abdomen of Drosophila melanogaster due to its prolongation of expression of the transcription factor Broad (BR). In a microarray analysis of JH-induced gene expression in abdominal integument, we found that Krüppel homolog 1 (Kr-h1) was up-regulated during most of adult development. Quantitative real-time PCR analyses showed that Kr-h1 up-regulation began at 10h after puparium formation (APF), and Kr-h1 up-regulation occurred in imaginal epidermal cells, persisting larval muscles, and larval oenocytes. Ectopic expression of Kr-h1 in abdominal epidermis using T155-Gal4 to drive UAS-Kr-h1 resulted in missing or short bristles in the dorsal midline. This phenotype was similar to that seen after a low dose of JH or after misexpression of br between 21 and 30 h APF. Ectopic expression of Kr-h1 prolonged the expression of BR protein in the pleura and the dorsal tergite. No Kr-h1 was seen after misexpression of br. Thus, Kr-h1 mediates some of the JH signaling in the adult abdominal epidermis and is upstream of br in this pathway. We also show for the first time that the JH-mediated maintenance of br expression in this epidermis is patterned and that JH delays the fusion of the imaginal cells and the disappearance of Dpp in the dorsal midline.
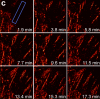
We demonstrate live-cell super-resolution imaging using photoactivated localization microscopy (PALM). The use of photon-tolerant cell lines in combination with the high resolution and molecular sensitivity of PALM permitted us to investigate the nanoscale dynamics within individual adhesion complexes (ACs) in living cells under physiological conditions for as long as 25 min, with half of the time spent collecting the PALM images at spatial resolutions down to approximately 60 nm and frame rates as short as 25 s. We visualized the formation of ACs and measured the fractional gain and loss of individual paxillin molecules as each AC evolved. By allowing observation of a wide variety of nanoscale dynamics, live-cell PALM provides insights into molecular assembly during the initiation, maturation and dissolution of cellular processes.
Commentary: The first example of true live cell and time lapse imaging by localization microscopy (as opposed to particle tracking), this paper uses the Nyquist criterion to establish a necessary condition for true spatial resolution based on the density of localized molecules – a condition often unmet in claims elsewhere in the superresolution literature.
By any method, higher spatiotemporal resolution requires increasing light exposure at the specimen, making noninvasive imaging increasingly difficult. Here, simultaneous differential interference contrast imaging is used to establish that cells behave physiologically before, during, and after PALM imaging. Similar controls are lacking from many supposed “live cell” superresolution demonstrations.
Time invariant description of synaptic connectivity in cortical circuits may be precluded by the ongoing growth and retraction of dendritic spines accompanied by the formation and elimination of synapses. On the other hand, the spatial arrangement of axonal and dendritic branches appears stable. This suggests that an invariant description of connectivity can be cast in terms of potential synapses, which are locations in the neuropil where an axon branch of one neuron is proximal to a dendritic branch of another neuron. In this paper, we attempt to reconstruct the potential connectivity in local cortical circuits of the cat primary visual cortex (V1). Based on multiple single-neuron reconstructions of axonal and dendritic arbors in 3 dimensions, we evaluate the expected number of potential synapses and the probability of potential connectivity among excitatory (pyramidal and spiny stellate) neurons and inhibitory basket cells. The results provide a quantitative description of structural organization of local cortical circuits. For excitatory neurons from different cortical layers, we compute local domains, which contain their potentially pre- and postsynaptic excitatory partners. These domains have columnar shapes with laminar specific radii and are roughly of the size of the ocular dominance column. Therefore, connections between most excitatory neurons in the ocular dominance column can be implemented by local synaptogenesis. Structural connectivity involving inhibitory basket cells is generally weaker than excitatory connectivity. Here, only nearby neurons are capable of establishing more than one potential synapse, implying that within the ocular dominance column these connections have more limited potential for circuit remodeling.
Several aspects of locomotor control have been ascribed to the central complex of the insect brain; however, the role of distinct substructures of this complex is not well known. The tay bridge1 (tay1) mutant of Drosophila melanogaster was originally isolated on the basis of reduced walking speed and activity. In addition, tay1 is defective in the compensation of rotatory stimuli during walking and histologically, tay1 causes a mid-sagittal constriction of the protocerebral bridge, a constituent of the central complex. Cloning of the tay gene revealed that it encodes a novel protein with no significant homology to any known protein. To associate the behavioral phenotypes with the anatomical defect in the protocerebral bridge, we used different driver lines to express the tay cDNA in various neuronal subpopulations of the central brain in tay1-mutant flies. These experiments showed an association of the aberrant walking speed and activity with the structural defect in the protocerebral bridge. In contrast, the compensation of rotatory stimuli during walking was rescued without a restoration of the protocerebral bridge. The results of our differential rescue approach are supported by neuronal silencing experiments using conditional tetanus toxin expression in the same subset of neurons. These findings show for the first time that the walking speed and activity is controlled by different substructures of the central brain than the compensatory locomotion for rotatory stimuli.
The mechanisms by which ethanol induces changes in behavior are not well understood. Here, we show that Caenorhabditis elegans loss-of-function mutations in the synaptic vesicle-associated RAB-3 protein and its guanosine triphosphate exchange factor AEX-3 confer resistance to the acute locomotor effects of ethanol. Similarly, mice lacking one or both copies of Rab3A are resistant to the ataxic and sedative effects of ethanol, and Rab3A haploinsufficiency increases voluntary ethanol consumption. These data suggest a conserved role of RAB-3-/RAB3A-regulated neurotransmitter release in ethanol-related behaviors.
We present a method for automatic full-precision alignment of the images in a tomographic tilt series. Full-precision automatic alignment of cryo electron microscopy images has remained a difficult challenge to date, due to the limited electron dose and low image contrast. These facts lead to poor signal to noise ratio (SNR) in the images, which causes automatic feature trackers to generate errors, even with high contrast gold particles as fiducial features. To enable fully automatic alignment for full-precision reconstructions, we frame the problem probabilistically as finding the most likely particle tracks given a set of noisy images, using contextual information to make the solution more robust to the noise in each image. To solve this maximum likelihood problem, we use Markov Random Fields (MRF) to establish the correspondence of features in alignment and robust optimization for projection model estimation. The resulting algorithm, called Robust Alignment and Projection Estimation for Tomographic Reconstruction, or RAPTOR, has not needed any manual intervention for the difficult datasets we have tried, and has provided sub-pixel alignment that is as good as the manual approach by an expert user. We are able to automatically map complete and partial marker trajectories and thus obtain highly accurate image alignment. Our method has been applied to challenging cryo electron tomographic datasets with low SNR from intact bacterial cells, as well as several plastic section and X-ray datasets.
The Honeybee Brain Atlas serves as 3D database and communicative platform to accumulate structural data, i.e. reconstructed neurons, derived from confocal scans (Brandt et al., 2005) (www.neurobiologie.fu-berlin.de/beebrain/) (1). Transforming neurons into the atlas requires manual segmentation of neuropils within confocal images, a time-consuming task requiring expertise in identifying biological structures which can result in different outcomes from various segmenters.
Meiosis is a highly conserved process in which a diploid genome is recombined and assorted into haploid gametes. Remarkably, the pea aphid Acyrthosiphon pisum exhibits a reproductive polyphenism whereby environmental signals trigger a switch between apomixis (parthenogenetic reproduction) and meiosis (sexual reproduction). Aphid apomixis results in daughter embryo clones with 2n genome content without male contribution or recombination. This important adaptation allows aphid populations to not only rapidly expand upon abundant resources during summer but also survive winter. How aphids have evolved this ability to switch between parthenogenesis and sexual meiosis is unknown. To arrive at a mechanistic explanation for this developmental plasticity, I determined meiosis gene activity in sexuals and asexuals. I first identified homologs of a core set of meiosis genes from the pea aphid genome. Next, I tested the expression of these core meiosis genes by PCR spanning across at least one intron from cDNA isolated from asexual and sexual ovaries. Surprisingly, meiosis specific genes (e.g., Spo11, Msh4, Msh5, Hop2 and Mnd1) are expressed in asexual ovaries. Additionally, the Spo11 PCR product contained intronic sequence, thus representing unspliced mRNA. Future experiments looking at the quantities and localizations of mRNA and protein will help to distinguish among several possible explanations for these results. Further molecular characterization of this phenotypic plasticity will be helpful in understanding how multiple interacting pathways can evolve to create alternate developmental phenotypes.