Filter
Associated Lab
- Aguilera Castrejon Lab (19) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (75) Apply Ahrens Lab filter
- Aso Lab (42) Apply Aso Lab filter
- Baker Lab (38) Apply Baker Lab filter
- Betzig Lab (116) Apply Betzig Lab filter
- Beyene Lab (15) Apply Beyene Lab filter
- Bock Lab (17) Apply Bock Lab filter
- Branson Lab (56) Apply Branson Lab filter
- Card Lab (43) Apply Card Lab filter
- Cardona Lab (64) Apply Cardona Lab filter
- Chklovskii Lab (13) Apply Chklovskii Lab filter
- Clapham Lab (16) Apply Clapham Lab filter
- Cui Lab (19) Apply Cui Lab filter
- Darshan Lab (12) Apply Darshan Lab filter
- Dennis Lab (3) Apply Dennis Lab filter
- Dickson Lab (46) Apply Dickson Lab filter
- Druckmann Lab (25) Apply Druckmann Lab filter
- Dudman Lab (58) Apply Dudman Lab filter
- Eddy/Rivas Lab (30) Apply Eddy/Rivas Lab filter
- Egnor Lab (11) Apply Egnor Lab filter
- Espinosa Medina Lab (25) Apply Espinosa Medina Lab filter
- Feliciano Lab (16) Apply Feliciano Lab filter
- Fetter Lab (41) Apply Fetter Lab filter
- FIB-SEM Technology (1) Apply FIB-SEM Technology filter
- Fitzgerald Lab (30) Apply Fitzgerald Lab filter
- Freeman Lab (15) Apply Freeman Lab filter
- Funke Lab (46) Apply Funke Lab filter
- Gonen Lab (91) Apply Gonen Lab filter
- Grigorieff Lab (62) Apply Grigorieff Lab filter
- Harris Lab (65) Apply Harris Lab filter
- Heberlein Lab (94) Apply Heberlein Lab filter
- Hermundstad Lab (32) Apply Hermundstad Lab filter
- Hess Lab (80) Apply Hess Lab filter
- Ilanges Lab (4) Apply Ilanges Lab filter
- Jayaraman Lab (49) Apply Jayaraman Lab filter
- Ji Lab (33) Apply Ji Lab filter
- Johnson Lab (7) Apply Johnson Lab filter
- Kainmueller Lab (19) Apply Kainmueller Lab filter
- Karpova Lab (15) Apply Karpova Lab filter
- Keleman Lab (13) Apply Keleman Lab filter
- Keller Lab (77) Apply Keller Lab filter
- Koay Lab (20) Apply Koay Lab filter
- Lavis Lab (162) Apply Lavis Lab filter
- Lee (Albert) Lab (34) Apply Lee (Albert) Lab filter
- Leonardo Lab (23) Apply Leonardo Lab filter
- Li Lab (32) Apply Li Lab filter
- Lippincott-Schwartz Lab (182) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (9) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (65) Apply Liu (Zhe) Lab filter
- Looger Lab (138) Apply Looger Lab filter
- Magee Lab (49) Apply Magee Lab filter
- Menon Lab (18) Apply Menon Lab filter
- Murphy Lab (13) Apply Murphy Lab filter
- O'Shea Lab (8) Apply O'Shea Lab filter
- Otopalik Lab (13) Apply Otopalik Lab filter
- Pachitariu Lab (56) Apply Pachitariu Lab filter
- Pastalkova Lab (19) Apply Pastalkova Lab filter
- Pavlopoulos Lab (19) Apply Pavlopoulos Lab filter
- Pedram Lab (15) Apply Pedram Lab filter
- Podgorski Lab (16) Apply Podgorski Lab filter
- Reiser Lab (55) Apply Reiser Lab filter
- Riddiford Lab (44) Apply Riddiford Lab filter
- Romani Lab (52) Apply Romani Lab filter
- Rubin Lab (149) Apply Rubin Lab filter
- Saalfeld Lab (66) Apply Saalfeld Lab filter
- Satou Lab (18) Apply Satou Lab filter
- Scheffer Lab (38) Apply Scheffer Lab filter
- Schreiter Lab (72) Apply Schreiter Lab filter
- Schulze Lab (1) Apply Schulze Lab filter
- Sgro Lab (23) Apply Sgro Lab filter
- Shroff Lab (35) Apply Shroff Lab filter
- Simpson Lab (23) Apply Simpson Lab filter
- Singer Lab (80) Apply Singer Lab filter
- Spruston Lab (98) Apply Spruston Lab filter
- Stern Lab (160) Apply Stern Lab filter
- Sternson Lab (54) Apply Sternson Lab filter
- Stringer Lab (44) Apply Stringer Lab filter
- Svoboda Lab (136) Apply Svoboda Lab filter
- Tebo Lab (36) Apply Tebo Lab filter
- Tervo Lab (10) Apply Tervo Lab filter
- Tillberg Lab (22) Apply Tillberg Lab filter
- Tjian Lab (64) Apply Tjian Lab filter
- Truman Lab (88) Apply Truman Lab filter
- Turaga Lab (53) Apply Turaga Lab filter
- Turner Lab (38) Apply Turner Lab filter
- Vale Lab (8) Apply Vale Lab filter
- Voigts Lab (5) Apply Voigts Lab filter
- Wang (Meng) Lab (31) Apply Wang (Meng) Lab filter
- Wang (Shaohe) Lab (25) Apply Wang (Shaohe) Lab filter
- Wong-Campos Lab (1) Apply Wong-Campos Lab filter
- Wu Lab (9) Apply Wu Lab filter
- Zlatic Lab (28) Apply Zlatic Lab filter
- Zuker Lab (25) Apply Zuker Lab filter
Associated Project Team
- CellMap (13) Apply CellMap filter
- COSEM (3) Apply COSEM filter
- FIB-SEM Technology (5) Apply FIB-SEM Technology filter
- Fly Descending Interneuron (12) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (14) Apply Fly Functional Connectome filter
- Fly Olympiad (5) Apply Fly Olympiad filter
- FlyEM (56) Apply FlyEM filter
- FlyLight (50) Apply FlyLight filter
- GENIE (47) Apply GENIE filter
- Integrative Imaging (11) Apply Integrative Imaging filter
- Larval Olympiad (2) Apply Larval Olympiad filter
- MouseLight (18) Apply MouseLight filter
- NeuroSeq (1) Apply NeuroSeq filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (29) Apply Tool Translation Team (T3) filter
- Transcription Imaging (49) Apply Transcription Imaging filter
Publication Date
- 2026 (118) Apply 2026 filter
- 2025 (223) Apply 2025 filter
- 2024 (209) Apply 2024 filter
- 2023 (158) Apply 2023 filter
- 2022 (192) Apply 2022 filter
- 2021 (194) Apply 2021 filter
- 2020 (196) Apply 2020 filter
- 2019 (202) Apply 2019 filter
- 2018 (232) Apply 2018 filter
- 2017 (217) Apply 2017 filter
- 2016 (209) Apply 2016 filter
- 2015 (252) Apply 2015 filter
- 2014 (236) Apply 2014 filter
- 2013 (194) Apply 2013 filter
- 2012 (190) Apply 2012 filter
- 2011 (190) Apply 2011 filter
- 2010 (161) Apply 2010 filter
- 2009 (158) Apply 2009 filter
- 2008 (140) Apply 2008 filter
- 2007 (106) Apply 2007 filter
- 2006 (92) Apply 2006 filter
- 2005 (67) Apply 2005 filter
- 2004 (57) Apply 2004 filter
- 2003 (58) Apply 2003 filter
- 2002 (39) Apply 2002 filter
- 2001 (28) Apply 2001 filter
- 2000 (29) Apply 2000 filter
- 1999 (14) Apply 1999 filter
- 1998 (18) Apply 1998 filter
- 1997 (16) Apply 1997 filter
- 1996 (10) Apply 1996 filter
- 1995 (18) Apply 1995 filter
- 1994 (12) Apply 1994 filter
- 1993 (10) Apply 1993 filter
- 1992 (6) Apply 1992 filter
- 1991 (11) Apply 1991 filter
- 1990 (11) Apply 1990 filter
- 1989 (6) Apply 1989 filter
- 1988 (1) Apply 1988 filter
- 1987 (7) Apply 1987 filter
- 1986 (4) Apply 1986 filter
- 1985 (5) Apply 1985 filter
- 1984 (2) Apply 1984 filter
- 1983 (2) Apply 1983 filter
- 1982 (3) Apply 1982 filter
- 1981 (3) Apply 1981 filter
- 1980 (1) Apply 1980 filter
- 1979 (1) Apply 1979 filter
- 1976 (2) Apply 1976 filter
- 1973 (1) Apply 1973 filter
- 1970 (1) Apply 1970 filter
- 1967 (1) Apply 1967 filter
Type of Publication
4313 Publications
Showing 531-540 of 4313 resultsArtificial activation of anatomically localized, genetically defined hypothalamic neuron populations is known to trigger distinct innate behaviors, suggesting a hypothalamic nucleus-centered organization of behavior control. To assess whether the encoding of behavior is similarly anatomically confined, we performed simultaneous neuron recordings across twenty hypothalamic regions in freely moving animals. Here we show that distinct but anatomically distributed neuron ensembles encode the social and fear behavior classes, primarily through mixed selectivity. While behavior class-encoding ensembles were spatially distributed, individual ensembles exhibited strong localization bias. Encoding models identified that behavior actions, but not motion-related variables, explained a large fraction of hypothalamic neuron activity variance. These results identify unexpected complexity in the hypothalamic encoding of instincts and provide a foundation for understanding the role of distributed neural representations in the expression of behaviors driven by hardwired circuits.
Vocal learning in songbirds requires an anatomically discrete and functionally dedicated circuit called the anterior forebrain pathway (AFP). The AFP is homologous to cortico-basal ganglia-thalamo-cortical loops in mammals. The basal ganglia portion of this pathway, Area X, shares many features characteristic of the mammalian striatum and pallidum, including cell types and connectivity. The AFP also deviates from mammalian basal ganglia circuits in fundamental ways. In addition, the microcircuitry, role of neuromodulators, and function of Area X are still unclear. Elucidating the mechanisms by which both mammalian-like and unique features of the AFP contribute to vocal learning may help lead to a broad understanding of the sensorimotor functions of basal ganglia circuits.
Intracellular recording allows precise measurement and manipulation of individual neurons, but it requires stable mechanical contact between the electrode and the cell membrane, and thus it has remained challenging to perform in behaving animals. Whole-cell recordings in freely moving animals can be obtained by rigidly fixing ('anchoring') the pipette electrode to the head; however, previous anchoring procedures were slow and often caused substantial pipette movement, resulting in loss of the recording or of recording quality. We describe a UV-transparent collar and UV-cured adhesive technique that rapidly (within 15 s) anchors pipettes in place with virtually no movement, thus substantially improving the reliability, yield and quality of freely moving whole-cell recordings. Recordings are first obtained from anesthetized or awake head-fixed rats. UV light cures the thin adhesive layers linking pipette to collar to head. Then, the animals are rapidly and smoothly released for recording during unrestrained behavior. The anesthetized-patched version can be completed in ∼4-7 h (excluding histology) and the awake-patched version requires ∼1-4 h per day for ∼2 weeks. These advances should greatly facilitate studies of neuronal integration and plasticity in identified cells during natural behaviors.
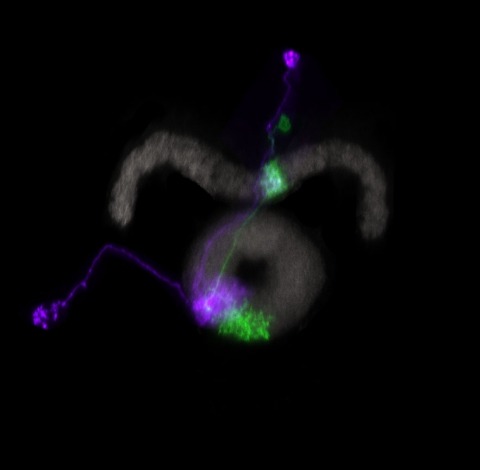
Many animals maintain an internal representation of their heading as they move through their surroundings. Such a compass representation was recently discovered in a neural population in the Drosophila melanogaster central complex, a brain region implicated in spatial navigation. Here, we use two-photon calcium imaging and electrophysiology in head-fixed walking flies to identify a different neural population that conjunctively encodes heading and angular velocity, and is excited selectively by turns in either the clockwise or counterclockwise direction. We show how these mirror-symmetric turn responses combine with the neurons' connectivity to the compass neurons to create an elegant mechanism for updating the fly's heading representation when the animal turns in darkness. This mechanism, which employs recurrent loops with an angular shift, bears a resemblance to those proposed in theoretical models for rodent head direction cells. Our results provide a striking example of structure matching function for a broadly relevant computation.
The field of connectomics has recently produced neuron wiring diagrams from relatively large brain regions from multiple animals. Most of these neural reconstructions were computed from isotropic (e.g., FIBSEM) or near isotropic (e.g., SBEM) data. In spite of the remarkable progress on algorithms in recent years, automatic dense reconstruction from anisotropic data remains a challenge for the connectomics community. One significant hurdle in the segmentation of anisotropic data is the difficulty in generating a suitable initial over-segmentation. In this study, we present a segmentation method for anisotropic EM data that agglomerates a 3D over-segmentation computed from the 3D affinity prediction. A 3D U-net is trained to predict 3D affinities by the MALIS approach. Experiments on multiple datasets demonstrates the strength and robustness of the proposed method for anisotropic EM segmentation.
Full reconstruction of neuron morphology is of fundamental interest for the analysis and understanding of their functioning. We have developed a novel method capable of automatically tracing neurons in three-dimensional microscopy data. In contrast to template-based methods, the proposed approach makes no assumptions about the shape or appearance of neurite structure. Instead, an efficient seeding approach is applied to capture complex neuronal structures and the tracing problem is solved by computing the optimal reconstruction with a weighted graph. The optimality is determined by the cost function designed for the path between each pair of seeds and by topological constraints defining the component interrelations and completeness. In addition, an automated neuron comparison method is introduced for performance evaluation and structure analysis. The proposed algorithm is computationally efficient and has been validated using different types of microscopy data sets including Drosophila’s projection neurons and fly neurons with presynaptic sites. In all cases, the approach yielded promising results.
How metazoan mechanotransduction channels sense mechanical stimuli is not well understood. The NOMPC channel in the transient receptor potential (TRP) family, a mechanotransduction channel for Drosophila touch sensation and hearing, contains 29 Ankyrin repeats (ARs) that associate with microtubules. These ARs have been postulated to act as a tether that conveys force to the channel. Here, we report that these N-terminal ARs form a cytoplasmic domain essential for NOMPC mechanogating in vitro, mechanosensitivity of touch receptor neurons in vivo, and touch-induced behaviors of Drosophila larvae. Duplicating the ARs elongates the filaments that tether NOMPC to microtubules in mechanosensory neurons. Moreover, microtubule association is required for NOMPC mechanogating. Importantly, transferring the NOMPC ARs to mechanoinsensitive voltage-gated potassium channels confers mechanosensitivity to the chimeric channels. These experiments strongly support a tether mechanism of mechanogating for the NOMPC channel, providing insights into the basis of mechanosensitivity of mechanotransduction channels.
Many different types of functional non-coding RNAs participate in a wide range of important cellular functions but the large majority of these RNAs are not routinely annotated in published genomes. Several programs have been developed for identifying RNAs, including specific tools tailored to a particular RNA family as well as more general ones designed to work for any family. Many of these tools utilize covariance models (CMs), statistical models of the conserved sequence, and structure of an RNA family. In this chapter, as an illustrative example, the Infernal software package and CMs from the Rfam database are used to identify RNAs in the genome of the archaeon Methanobrevibacter ruminantium, uncovering some additional RNAs not present in the genome’s initial annotation. Analysis of the results and comparison with family-specific methods demonstrate some important strengths and weaknesses of this general approach.
Reconstructing neuronal circuits at the level of synapses is a central problem in neuroscience and becoming a focus of the emerging field of connectomics. To date, electron microscopy (EM) is the most proven technique for identifying and quantifying synaptic connections. As advances in EM make acquiring larger datasets possible, subsequent manual synapse identification ({\em i.e.}, proofreading) for deciphering a connectome becomes a major time bottleneck. Here we introduce a large-scale, high-throughput, and semi-automated methodology to efficiently identify synapses. We successfully applied our methodology to the Drosophila medulla optic lobe, annotating many more synapses than previous connectome efforts. Our approaches are extensible and will make the often complicated process of synapse identification accessible to a wider-community of potential proofreaders.
The anterolateral motor cortex (ALM) and ventromedial (VM) thalamus are functionally linked to support persistent activity during motor planning. We analyzed the underlying synaptic interconnections using optogenetics and electrophysiology in mice (♀/♂). In cortex, thalamocortical (TC) axons from VM excited VM-projecting pyramidal-tract (PT) neurons in layer 5B of ALM. These axons also strongly excited layer 2/3 neurons (which strongly excite PT neurons, as previously shown) but not VM-projecting corticothalamic (CT) neurons in layer 6. The strongest connections in the VM→PT circuit were localized to apical-tuft dendrites of PT neurons, in layer 1. These tuft inputs were selectively augmented after blocking hyperpolarization-activated cyclic nucleotide-gated (HCN) channels. In thalamus, axons from ALM PT neurons excited ALM-projecting VM neurons, located medially in VM. These axons provided weak input to neurons in mediodorsal nucleus, and little or no input either to neurons in the GABAergic reticular thalamic nucleus or to neurons in VM projecting to primary motor cortex (M1). Conversely, M1 PT axons excited M1- but not ALM-projecting VM neurons. Our findings indicate, first, a set of cell-type-specific connections forming an excitatory thalamo-cortico-thalamic (T-C-T) loop for ALM↔VM communication and a circuit-level substrate for supporting reverberant activity in this system. Second, a key feature of this loop is the prominent involvement of layer 1 synapses onto apical dendrites, a subcellular compartment with distinct signaling properties, including HCN-mediated gain control. Third, the segregation of the ALM↔VM loop from M1-related circuits of VM adds cellular-level support for the concept of parallel pathway organization in the motor system.Anterolateral motor cortex (ALM), a higher-order motor area in the mouse, and ventromedial thalamus (VM) are anatomically and functionally linked, but their synaptic interconnections at the cellular level are unknown. Our results show that ALM pyramidal tract neurons monosynaptically excite ALM-projecting thalamocortical neurons in a medial subdivision of VM, and vice versa. The thalamo-cortico-thalamic loop formed by these recurrent connections constitutes a circuit-level substrate for supporting reverberant activity in this system.