Filter
Associated Lab
- Ahrens Lab (3) Apply Ahrens Lab filter
- Aso Lab (6) Apply Aso Lab filter
- Baker Lab (2) Apply Baker Lab filter
- Betzig Lab (11) Apply Betzig Lab filter
- Branson Lab (6) Apply Branson Lab filter
- Cardona Lab (5) Apply Cardona Lab filter
- Chklovskii Lab (2) Apply Chklovskii Lab filter
- Cui Lab (5) Apply Cui Lab filter
- Dickson Lab (1) Apply Dickson Lab filter
- Druckmann Lab (3) Apply Druckmann Lab filter
- Dudman Lab (3) Apply Dudman Lab filter
- Eddy/Rivas Lab (4) Apply Eddy/Rivas Lab filter
- Egnor Lab (1) Apply Egnor Lab filter
- Fetter Lab (5) Apply Fetter Lab filter
- Freeman Lab (7) Apply Freeman Lab filter
- Funke Lab (1) Apply Funke Lab filter
- Gonen Lab (5) Apply Gonen Lab filter
- Grigorieff Lab (7) Apply Grigorieff Lab filter
- Harris Lab (7) Apply Harris Lab filter
- Heberlein Lab (1) Apply Heberlein Lab filter
- Hess Lab (7) Apply Hess Lab filter
- Jayaraman Lab (4) Apply Jayaraman Lab filter
- Ji Lab (4) Apply Ji Lab filter
- Karpova Lab (1) Apply Karpova Lab filter
- Keleman Lab (2) Apply Keleman Lab filter
- Keller Lab (7) Apply Keller Lab filter
- Lavis Lab (5) Apply Lavis Lab filter
- Leonardo Lab (2) Apply Leonardo Lab filter
- Liu (Zhe) Lab (4) Apply Liu (Zhe) Lab filter
- Looger Lab (9) Apply Looger Lab filter
- Magee Lab (5) Apply Magee Lab filter
- Murphy Lab (1) Apply Murphy Lab filter
- Pastalkova Lab (3) Apply Pastalkova Lab filter
- Reiser Lab (2) Apply Reiser Lab filter
- Romani Lab (2) Apply Romani Lab filter
- Rubin Lab (16) Apply Rubin Lab filter
- Saalfeld Lab (3) Apply Saalfeld Lab filter
- Scheffer Lab (2) Apply Scheffer Lab filter
- Schreiter Lab (4) Apply Schreiter Lab filter
- Shroff Lab (1) Apply Shroff Lab filter
- Simpson Lab (4) Apply Simpson Lab filter
- Singer Lab (10) Apply Singer Lab filter
- Spruston Lab (7) Apply Spruston Lab filter
- Stern Lab (4) Apply Stern Lab filter
- Sternson Lab (7) Apply Sternson Lab filter
- Svoboda Lab (9) Apply Svoboda Lab filter
- Tjian Lab (6) Apply Tjian Lab filter
- Truman Lab (6) Apply Truman Lab filter
- Turaga Lab (1) Apply Turaga Lab filter
- Turner Lab (2) Apply Turner Lab filter
- Wu Lab (1) Apply Wu Lab filter
- Zlatic Lab (4) Apply Zlatic Lab filter
- Zuker Lab (2) Apply Zuker Lab filter
Associated Project Team
Associated Support Team
- Electron Microscopy (1) Apply Electron Microscopy filter
- Gene Targeting and Transgenics (3) Apply Gene Targeting and Transgenics filter
- Invertebrate Shared Resource (1) Apply Invertebrate Shared Resource filter
- Janelia Experimental Technology (2) Apply Janelia Experimental Technology filter
- Management Team (1) Apply Management Team filter
- Molecular Genomics (1) Apply Molecular Genomics filter
- Primary & iPS Cell Culture (2) Apply Primary & iPS Cell Culture filter
- Project Technical Resources (1) Apply Project Technical Resources filter
- Scientific Computing Software (11) Apply Scientific Computing Software filter
Publication Date
- December 2015 (15) Apply December 2015 filter
- November 2015 (22) Apply November 2015 filter
- October 2015 (16) Apply October 2015 filter
- September 2015 (16) Apply September 2015 filter
- August 2015 (17) Apply August 2015 filter
- July 2015 (18) Apply July 2015 filter
- June 2015 (16) Apply June 2015 filter
- May 2015 (16) Apply May 2015 filter
- April 2015 (18) Apply April 2015 filter
- March 2015 (16) Apply March 2015 filter
- February 2015 (15) Apply February 2015 filter
- January 2015 (10) Apply January 2015 filter
- Remove 2015 filter 2015
195 Janelia Publications
Showing 191-195 of 195 resultsIn animals, Hox transcription factors define regional identity in distinct anatomical domains. How Hox genes encode this specificity is a paradox, because different Hox proteins bind with high affinity in vitro to similar DNA sequences. Here, we demonstrate that the Hox protein Ultrabithorax (Ubx) in complex with its cofactor Extradenticle (Exd) bound specifically to clusters of very low affinity sites in enhancers of the shavenbaby gene of Drosophila. These low affinity sites conferred specificity for Ubx binding in vivo, but multiple clustered sites were required for robust expression when embryos developed in variable environments. Although most individual Ubx binding sites are not evolutionarily conserved, the overall enhancer architecture-clusters of low affinity binding sites-is maintained and required for enhancer function. Natural selection therefore works at the level of the enhancer, requiring a particular density of low affinity Ubx sites to confer both specific and robust expression.
The mammalian taste system is responsible for sensing and responding to the five basic taste qualities: sweet, sour, bitter, salty and umami. Previously, we showed that each taste is detected by dedicated taste receptor cells (TRCs) on the tongue and palate epithelium. To understand how TRCs transmit information to higher neural centres, we examined the tuning properties of large ensembles of neurons in the first neural station of the gustatory system. Here, we generated and characterized a collection of transgenic mice expressing a genetically encoded calcium indicator in central and peripheral neurons, and used a gradient refractive index microendoscope combined with high-resolution two-photon microscopy to image taste responses from ganglion neurons buried deep at the base of the brain. Our results reveal fine selectivity in the taste preference of ganglion neurons; demonstrate a strong match between TRCs in the tongue and the principal neural afferents relaying taste information to the brain; and expose the highly specific transfer of taste information between taste cells and the central nervous system.
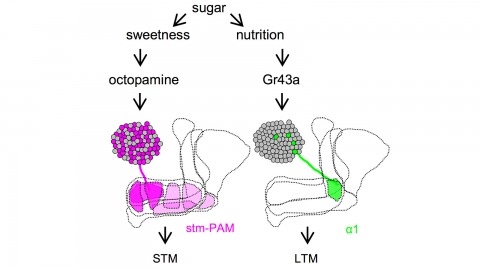
Drosophila melanogaster can acquire a stable appetitive olfactory memory when the presentation of a sugar reward and an odor are paired. However, the neuronal mechanisms by which a single training induces long-term memory are poorly understood. Here we show that two distinct subsets of dopamine neurons in the fly brain signal reward for short-term (STM) and long-term memories (LTM). One subset induces memory that decays within several hours, whereas the other induces memory that gradually develops after training. They convey reward signals to spatially segregated synaptic domains of the mushroom body (MB), a potential site for convergence. Furthermore, we identified a single type of dopamine neuron that conveys the reward signal to restricted subdomains of the mushroom body lobes and induces long-term memory. Constant appetitive memory retention after a single training session thus comprises two memory components triggered by distinct dopamine neurons.
Complex animal behaviors are built from dynamical relationships between sensory inputs, neuronal activity, and motor outputs in patterns with strategic value. Connecting these patterns illuminates how nervous systems compute behavior. Here, we study Drosophila larva navigation up temperature gradients toward preferred temperatures (positive thermotaxis). By tracking the movements of animals responding to fixed spatial temperature gradients or random temperature fluctuations, we calculate the sensitivity and dynamics of the conversion of thermosensory inputs into motor responses. We discover three thermosensory neurons in each dorsal organ ganglion (DOG) that are required for positive thermotaxis. Random optogenetic stimulation of the DOG thermosensory neurons evokes behavioral patterns that mimic the response to temperature variations. In vivo calcium and voltage imaging reveals that the DOG thermosensory neurons exhibit activity patterns with sensitivity and dynamics matched to the behavioral response. Temporal processing of temperature variations carried out by the DOG thermosensory neurons emerges in distinct motor responses during thermotaxis.
Tsetse flies (Glossina spp.), vectors of African trypanosomes, are distinguished by their specialized reproductive biology, defined by adenotrophic viviparity (maternal nourishment of progeny by glandular secretions followed by live birth). This trait has evolved infrequently among insects and requires unique reproductive mechanisms. A key event in Glossina reproduction involves the transition between periods of lactation and nonlactation (dry periods). Increased lipolysis, nutrient transfer to the milk gland, and milk-specific protein production characterize lactation, which terminates at the birth of the progeny and is followed by a period of involution. The dry stage coincides with embryogenesis of the progeny, during which lipid reserves accumulate in preparation for the next round of lactation. The obligate bacterial symbiont Wigglesworthia glossinidia is critical to tsetse reproduction and likely provides B vitamins required for metabolic processes underlying lactation and/or progeny development. Here we describe findings that utilized transcriptomics, physiological assays, and RNA interference-based functional analysis to understand different components of adenotrophic viviparity in tsetse flies.