Filter
Associated Lab
- Ahrens Lab (3) Apply Ahrens Lab filter
- Aso Lab (3) Apply Aso Lab filter
- Baker Lab (5) Apply Baker Lab filter
- Betzig Lab (8) Apply Betzig Lab filter
- Branson Lab (6) Apply Branson Lab filter
- Card Lab (1) Apply Card Lab filter
- Cardona Lab (1) Apply Cardona Lab filter
- Chklovskii Lab (2) Apply Chklovskii Lab filter
- Cui Lab (2) Apply Cui Lab filter
- Dickson Lab (5) Apply Dickson Lab filter
- Druckmann Lab (1) Apply Druckmann Lab filter
- Dudman Lab (4) Apply Dudman Lab filter
- Eddy/Rivas Lab (3) Apply Eddy/Rivas Lab filter
- Egnor Lab (1) Apply Egnor Lab filter
- Fetter Lab (1) Apply Fetter Lab filter
- Freeman Lab (3) Apply Freeman Lab filter
- Gonen Lab (10) Apply Gonen Lab filter
- Grigorieff Lab (3) Apply Grigorieff Lab filter
- Harris Lab (2) Apply Harris Lab filter
- Heberlein Lab (1) Apply Heberlein Lab filter
- Hess Lab (5) Apply Hess Lab filter
- Jayaraman Lab (1) Apply Jayaraman Lab filter
- Ji Lab (3) Apply Ji Lab filter
- Karpova Lab (1) Apply Karpova Lab filter
- Keller Lab (7) Apply Keller Lab filter
- Lavis Lab (7) Apply Lavis Lab filter
- Lee (Albert) Lab (4) Apply Lee (Albert) Lab filter
- Leonardo Lab (4) Apply Leonardo Lab filter
- Liu (Zhe) Lab (4) Apply Liu (Zhe) Lab filter
- Looger Lab (11) Apply Looger Lab filter
- Magee Lab (1) Apply Magee Lab filter
- Menon Lab (3) Apply Menon Lab filter
- Murphy Lab (1) Apply Murphy Lab filter
- Reiser Lab (2) Apply Reiser Lab filter
- Riddiford Lab (4) Apply Riddiford Lab filter
- Rubin Lab (8) Apply Rubin Lab filter
- Saalfeld Lab (1) Apply Saalfeld Lab filter
- Scheffer Lab (7) Apply Scheffer Lab filter
- Simpson Lab (2) Apply Simpson Lab filter
- Singer Lab (4) Apply Singer Lab filter
- Spruston Lab (1) Apply Spruston Lab filter
- Stern Lab (6) Apply Stern Lab filter
- Sternson Lab (5) Apply Sternson Lab filter
- Svoboda Lab (7) Apply Svoboda Lab filter
- Tervo Lab (1) Apply Tervo Lab filter
- Tjian Lab (4) Apply Tjian Lab filter
- Truman Lab (1) Apply Truman Lab filter
- Wu Lab (2) Apply Wu Lab filter
- Zlatic Lab (1) Apply Zlatic Lab filter
- Zuker Lab (1) Apply Zuker Lab filter
Associated Project Team
Associated Support Team
Publication Date
- December 2014 (31) Apply December 2014 filter
- November 2014 (7) Apply November 2014 filter
- October 2014 (14) Apply October 2014 filter
- September 2014 (14) Apply September 2014 filter
- August 2014 (11) Apply August 2014 filter
- July 2014 (23) Apply July 2014 filter
- June 2014 (13) Apply June 2014 filter
- May 2014 (10) Apply May 2014 filter
- April 2014 (18) Apply April 2014 filter
- March 2014 (13) Apply March 2014 filter
- February 2014 (11) Apply February 2014 filter
- January 2014 (25) Apply January 2014 filter
- Remove 2014 filter 2014
190 Janelia Publications
Showing 51-60 of 190 resultsThe superficial superior colliculus (sSC) occupies a critical node in the mammalian visual system; it is one of two major retinorecipient areas, receives visual cortical input, and innervates visual thalamocortical circuits. Nonetheless, the contribution of sSC neurons to downstream neural activity and visually guided behavior is unknown and frequently neglected. Here we identified the visual stimuli to which specific classes of sSC neurons respond, the downstream regions they target, and transgenic mice enabling class-specific manipulations. One class responds to small, slowly moving stimuli and projects exclusively to lateral posterior thalamus; another, comprising GABAergic neurons, responds to the sudden appearance or rapid movement of large stimuli and projects to multiple areas, including the lateral geniculate nucleus. A third class exhibits direction-selective responses and targets deeper SC layers. Together, our results show how specific sSC neurons represent and distribute diverse information and enable direct tests of their functional role.
Mitochondria are complex organelles at the center of cellular metabolism, apoptosis, and signaling. They continue to be the subject of intense basic investigation to understand their composition and function, but they have also captivated the attention of clinical researchers because of the growing knowledge of the (sometimes unexpected) roles of mitochondria in human diseases and aging. A full understanding of these intriguing organelles often requires their purification from cells or tissues under specific physiological or pathological conditions. Here we provide some introductory considerations for those interested in purifying mitochondria for subsequent downstream biophysical, structural, and functional analysis.
Motor sequences are formed through the serial execution of different movements, but how nervous systems implement this process remains largely unknown. We determined the organizational principles governing how dirty fruit flies groom their bodies with sequential movements. Using genetically targeted activation of neural subsets, we drove distinct motor programs that clean individual body parts. This enabled competition experiments revealing that the motor programs are organized into a suppression hierarchy; motor programs that occur first suppress those that occur later. Cleaning one body part reduces the sensory drive to its motor program, which relieves suppression of the next movement, allowing the grooming sequence to progress down the hierarchy. A model featuring independently evoked cleaning movements activated in parallel, but selected serially through hierarchical suppression, was successful in reproducing the grooming sequence. This provides the first example of an innate motor sequence implemented by the prevailing model for generating human action sequences.
A major limitation in understanding embryonic development is the lack of cell type-specific markers. Existing gene expression and marker atlases provide valuable tools, but they typically have one or more limitations: a lack of single-cell resolution; an inability to register multiple expression patterns to determine their precise relationship; an inability to be upgraded by users; an inability to compare novel patterns with the database patterns; and a lack of three-dimensional images. Here, we develop new 'atlas-builder' software that overcomes each of these limitations. A newly generated atlas is three-dimensional, allows the precise registration of an infinite number of cell type-specific markers, is searchable and is open-ended. Our software can be used to create an atlas of any tissue in any organism that contains stereotyped cell positions. We used the software to generate an 'eNeuro' atlas of the Drosophila embryonic CNS containing eight transcription factors that mark the major CNS cell types (motor neurons, glia, neurosecretory cells and interneurons). We found neuronal, but not glial, nuclei occupied stereotyped locations. We added 75 new Gal4 markers to the atlas to identify over 50% of all interneurons in the ventral CNS, and these lines allowed functional access to those interneurons for the first time. We expect the atlas-builder software to benefit a large proportion of the developmental biology community, and the eNeuro atlas to serve as a publicly accessible hub for integrating neuronal attributes - cell lineage, gene expression patterns, axon/dendrite projections, neurotransmitters--and linking them to individual neurons.
Distal enhancers commonly contact target promoters via chromatin looping. In erythroid cells, the locus control region (LCR) contacts β-type globin genes in a developmental stage-specific manner to stimulate transcription. Previously, we induced LCR-promoter looping by tethering the self-association domain (SA) of Ldb1 to the β-globin promoter via artificial zinc fingers. Here, we show that targeting the SA to a developmentally silenced embryonic globin gene in adult murine erythroblasts triggers its transcriptional reactivation. This activity depends on the LCR, consistent with an LCR-promoter looping mechanism. Strikingly, targeting the SA to the fetal γ-globin promoter in primary adult human erythroblasts increases γ-globin promoter-LCR contacts, stimulating transcription to approximately 85% of total β-globin synthesis, with a reciprocal reduction in adult β-globin expression. Our findings demonstrate that forced chromatin looping can override a stringent developmental gene expression program and suggest a novel approach to control the balance of globin gene transcription for therapeutic applications.
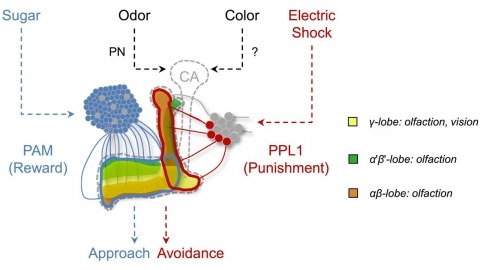
In nature, animals form memories associating reward or punishment with stimuli from different sensory modalities, such as smells and colors. It is unclear, however, how distinct sensory memories are processed in the brain. We established appetitive and aversive visual learning assays for Drosophila that are comparable to the widely used olfactory learning assays. These assays share critical features, such as reinforcing stimuli (sugar reward and electric shock punishment), and allow direct comparison of the cellular requirements for visual and olfactory memories. We found that the same subsets of dopamine neurons drive formation of both sensory memories. Furthermore, distinct yet partially overlapping subsets of mushroom body intrinsic neurons are required for visual and olfactory memories. Thus, our results suggest that distinct sensory memories are processed in a common brain center. Such centralization of related brain functions is an economical design that avoids the repetition of similar circuit motifs.
Genetically-encoded calcium indicators (GECIs) facilitate imaging activity of genetically defined neuronal populations in vivo. The high intracellular GECI concentrations required for in vivo imaging are usually achieved by viral gene transfer using adeno-associated viruses. Transgenic expression of GECIs promises important advantages, including homogeneous, repeatable, and stable expression without the need for invasive virus injections. Here we present the generation and characterization of transgenic mice expressing the GECIs GCaMP6s or GCaMP6f under the Thy1 promoter. We quantified GCaMP6 expression across brain regions and neurons and compared to other transgenic mice and AAV-mediated expression. We tested three mouse lines for imaging in the visual cortex in vivo and compared their performance to mice injected with AAV expressing GCaMP6. Furthermore, we show that GCaMP6 Thy1 transgenic mice are useful for long-term, high-sensitivity imaging in behaving mice.
Advances in chemistry and physics have profound effects on neuroimaging. Current and future progress in these disciplines will continue to aid in efforts to visualize neural circuitry, particularly in deeper layers of the brain.
Pixel and superpixel classifiers have become essential tools for EM segmentation algorithms. Training these classifiers remains a major bottleneck primarily due to the requirement of completely annotating the dataset which is tedious, error-prone and costly. In this paper, we propose an interactive learning scheme for the superpixel classifier for EM segmentation. Our algorithm is 'active semi-supervised' because it requests the labels of a small number of examples from user and applies label propagation technique to generate these queries. Using only a small set (< 20%) of all datapoints, the proposed algorithm consistently generates a classifier almost as accurate as that estimated from a complete groundtruth. We provide segmentation results on multiple datasets to show the strength of these classifiers.

The origin of chordates has been debated for more than a century, with one key issue being the emergence of the notochord. In vertebrates, the notochord develops by convergence and extension of the chordamesoderm, a population of midline cells of unique molecular identity. We identify a population of mesodermal cells in a developing invertebrate, the marine annelid Platynereis dumerilii, that converges and extends toward the midline and expresses a notochord-specific combination of genes. These cells differentiate into a longitudinal muscle, the axochord, that is positioned between central nervous system and axial blood vessel and secretes a strong collagenous extracellular matrix. Ancestral state reconstruction suggests that contractile mesodermal midline cells existed in bilaterian ancestors. We propose that these cells, via vacuolization and stiffening, gave rise to the chordate notochord.