Filter
Result Type
- Apply filter
- Apply filter
- Apply filter
- Apply filter
- Apply filter
- Apply filter
- Apply filter
- Apply filter
- Apply filter
- Area Landing Page (8) Apply Area Landing Page filter
- Collaborations (2) Apply Collaborations filter
- Conferences (241) Apply Conferences filter
- Janelia Archives (19) Apply Janelia Archives filter
- Janelia Archives Landing (1) Apply Janelia Archives Landing filter
- Lab (59) Apply Lab filter
- News Stories (270) Apply News Stories filter
- Other (543) Apply Other filter
- People (647) Apply People filter
- Project Team (15) Apply Project Team filter
- Publications (2647) Apply Publications filter
- Support Team (21) Apply Support Team filter
- Theory Fellow Landing Page (4) Apply Theory Fellow Landing Page filter
- Tool (128) Apply Tool filter
Associated Lab
- Aguilera Castrejon Lab (6) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (71) Apply Ahrens Lab filter
- Aso Lab (48) Apply Aso Lab filter
- Baker Lab (20) Apply Baker Lab filter
- Betzig Lab (111) Apply Betzig Lab filter
- Beyene Lab (14) Apply Beyene Lab filter
- Bock Lab (15) Apply Bock Lab filter
- Branson Lab (59) Apply Branson Lab filter
- Card Lab (35) Apply Card Lab filter
- Cardona Lab (44) Apply Cardona Lab filter
- Chklovskii Lab (10) Apply Chklovskii Lab filter
- Clapham Lab (23) Apply Clapham Lab filter
- Cui Lab (20) Apply Cui Lab filter
- Darshan Lab (8) Apply Darshan Lab filter
- Dennis Lab (7) Apply Dennis Lab filter
- Dickson Lab (34) Apply Dickson Lab filter
- Druckmann Lab (21) Apply Druckmann Lab filter
- Dudman Lab (50) Apply Dudman Lab filter
- Eddy/Rivas Lab (30) Apply Eddy/Rivas Lab filter
- Egnor Lab (5) Apply Egnor Lab filter
- Espinosa Medina Lab (25) Apply Espinosa Medina Lab filter
- Feliciano Lab (14) Apply Feliciano Lab filter
- Fetter Lab (31) Apply Fetter Lab filter
- Fitzgerald Lab (16) Apply Fitzgerald Lab filter
- Freeman Lab (16) Apply Freeman Lab filter
- Funke Lab (45) Apply Funke Lab filter
- Gonen Lab (60) Apply Gonen Lab filter
- Grigorieff Lab (34) Apply Grigorieff Lab filter
- Harris Lab (62) Apply Harris Lab filter
- Heberlein Lab (15) Apply Heberlein Lab filter
- Hermundstad Lab (30) Apply Hermundstad Lab filter
- Hess Lab (84) Apply Hess Lab filter
- Ilanges Lab (9) Apply Ilanges Lab filter
- Jayaraman Lab (57) Apply Jayaraman Lab filter
- Ji Lab (34) Apply Ji Lab filter
- Johnson Lab (8) Apply Johnson Lab filter
- Kainmueller Lab (1) Apply Kainmueller Lab filter
- Karpova Lab (24) Apply Karpova Lab filter
- Keleman Lab (8) Apply Keleman Lab filter
- Keller Lab (81) Apply Keller Lab filter
- Koay Lab (8) Apply Koay Lab filter
- Lavis Lab (155) Apply Lavis Lab filter
- Lee (Albert) Lab (32) Apply Lee (Albert) Lab filter
- Leonardo Lab (19) Apply Leonardo Lab filter
- Li Lab (11) Apply Li Lab filter
- Lippincott-Schwartz Lab (108) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (8) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (64) Apply Liu (Zhe) Lab filter
- Looger Lab (143) Apply Looger Lab filter
- Magee Lab (31) Apply Magee Lab filter
- Menon Lab (12) Apply Menon Lab filter
- Murphy Lab (7) Apply Murphy Lab filter
- O'Shea Lab (12) Apply O'Shea Lab filter
- Otopalik Lab (9) Apply Otopalik Lab filter
- Pachitariu Lab (42) Apply Pachitariu Lab filter
- Pastalkova Lab (6) Apply Pastalkova Lab filter
- Pavlopoulos Lab (7) Apply Pavlopoulos Lab filter
- Pedram Lab (12) Apply Pedram Lab filter
- Podgorski Lab (19) Apply Podgorski Lab filter
- Reiser Lab (62) Apply Reiser Lab filter
- Riddiford Lab (21) Apply Riddiford Lab filter
- Romani Lab (45) Apply Romani Lab filter
- Rubin Lab (123) Apply Rubin Lab filter
- Ryan Lab (1) Apply Ryan Lab filter
- Saalfeld Lab (57) Apply Saalfeld Lab filter
- Satou Lab (8) Apply Satou Lab filter
- Scheffer Lab (39) Apply Scheffer Lab filter
- Schreiter Lab (62) Apply Schreiter Lab filter
- Sgro Lab (10) Apply Sgro Lab filter
- Shroff Lab (43) Apply Shroff Lab filter
- Simpson Lab (18) Apply Simpson Lab filter
- Singer Lab (39) Apply Singer Lab filter
- Spruston Lab (75) Apply Spruston Lab filter
- Stern Lab (83) Apply Stern Lab filter
- Sternson Lab (52) Apply Sternson Lab filter
- Stringer Lab (37) Apply Stringer Lab filter
- Svoboda Lab (145) Apply Svoboda Lab filter
- Tebo Lab (20) Apply Tebo Lab filter
- Tervo Lab (14) Apply Tervo Lab filter
- Tillberg Lab (22) Apply Tillberg Lab filter
- Tjian Lab (19) Apply Tjian Lab filter
- Truman Lab (59) Apply Truman Lab filter
- Turaga Lab (52) Apply Turaga Lab filter
- Turner Lab (33) Apply Turner Lab filter
- Vale Lab (13) Apply Vale Lab filter
- Voigts Lab (9) Apply Voigts Lab filter
- Wang (Meng) Lab (31) Apply Wang (Meng) Lab filter
- Wang (Shaohe) Lab (11) Apply Wang (Shaohe) Lab filter
- Wong-Campos Lab (4) Apply Wong-Campos Lab filter
- Wu Lab (9) Apply Wu Lab filter
- Zlatic Lab (26) Apply Zlatic Lab filter
- Zuker Lab (5) Apply Zuker Lab filter
Associated Project Team
- CellMap (39) Apply CellMap filter
- COSEM (3) Apply COSEM filter
- FIB-SEM Technology (8) Apply FIB-SEM Technology filter
- Fly Descending Interneuron (12) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (15) Apply Fly Functional Connectome filter
- Fly Olympiad (5) Apply Fly Olympiad filter
- FlyEM (64) Apply FlyEM filter
- FlyLight (58) Apply FlyLight filter
- GENIE (67) Apply GENIE filter
- Integrative Imaging (2) Apply Integrative Imaging filter
- Larval Olympiad (2) Apply Larval Olympiad filter
- MouseLight (26) Apply MouseLight filter
- NeuroSeq (2) Apply NeuroSeq filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (36) Apply Tool Translation Team (T3) filter
- Transcription Imaging (48) Apply Transcription Imaging filter
Associated Support Team
- Project Pipeline Support (14) Apply Project Pipeline Support filter
- Anatomy and Histology (24) Apply Anatomy and Histology filter
- Cryo-Electron Microscopy (41) Apply Cryo-Electron Microscopy filter
- Electron Microscopy (21) Apply Electron Microscopy filter
- Flow Cytometry (4) Apply Flow Cytometry filter
- Gene Targeting and Transgenics (19) Apply Gene Targeting and Transgenics filter
- Immortalized Cell Line Culture (6) Apply Immortalized Cell Line Culture filter
- Integrative Imaging (32) Apply Integrative Imaging filter
- Invertebrate Shared Resource (50) Apply Invertebrate Shared Resource filter
- Janelia Experimental Technology (103) Apply Janelia Experimental Technology filter
- Management Team (1) Apply Management Team filter
- Mass Spectrometry (4) Apply Mass Spectrometry filter
- Media Facil\ (6) Apply Media Facil\ filter
- Molecular Genomics (22) Apply Molecular Genomics filter
- Primary & iPS Cell Culture (24) Apply Primary & iPS Cell Culture filter
- Project Technical Resources (61) Apply Project Technical Resources filter
- Quantitative Genomics (26) Apply Quantitative Genomics filter
- Scientific Computing Software (128) Apply Scientific Computing Software filter
- Scientific Computing Systems (13) Apply Scientific Computing Systems filter
- Viral Tools (22) Apply Viral Tools filter
- Vivarium (10) Apply Vivarium filter
Publication Date
- 2025 (85) Apply 2025 filter
- 2024 (252) Apply 2024 filter
- 2023 (192) Apply 2023 filter
- 2022 (193) Apply 2022 filter
- 2021 (187) Apply 2021 filter
- 2020 (194) Apply 2020 filter
- 2019 (201) Apply 2019 filter
- 2018 (221) Apply 2018 filter
- 2017 (202) Apply 2017 filter
- 2016 (207) Apply 2016 filter
- 2015 (222) Apply 2015 filter
- 2014 (216) Apply 2014 filter
- 2013 (152) Apply 2013 filter
- 2012 (112) Apply 2012 filter
- 2011 (98) Apply 2011 filter
- 2010 (61) Apply 2010 filter
- 2009 (56) Apply 2009 filter
- 2008 (40) Apply 2008 filter
- 2007 (21) Apply 2007 filter
- 2006 (3) Apply 2006 filter
Tool Types
- Data (9) Apply Data filter
- Data Application (7) Apply Data Application filter
- Figshare (1) Apply Figshare filter
- Human Health (2) Apply Human Health filter
- Imaging Instrumentation (11) Apply Imaging Instrumentation filter
- Laboratory Hardware (3) Apply Laboratory Hardware filter
- Laboratory Tool (6) Apply Laboratory Tool filter
- Laboratory Tools (51) Apply Laboratory Tools filter
- Medical Technology (1) Apply Medical Technology filter
- Model Organisms (9) Apply Model Organisms filter
- Reagents (25) Apply Reagents filter
- Software (20) Apply Software filter
4761 Results
Showing 121-130 of 4761 resultsWe report the larval CNS expression patterns for 6,650 GAL4 lines based on cis-regulatory regions (CRMs) from the Drosophila genome. Adult CNS expression patterns were previously reported for this collection, thereby providing a unique resource for determining the origins of adult cells. An illustrative example reveals the origin of the astrocyte-like glia of the ventral CNS. Besides larval neurons and glia, the larval CNS contains scattered lineages of immature, adult-specific neurons. Comparison of lineage expression within this large collection of CRMs provides insight into the codes used for designating neuronal types. The CRMs encode both dense and sparse patterns of lineage expression. There is little correlation between brain and thoracic lineages in patterns of sparse expression, but expression in the two regions is highly correlated in the dense mode. The optic lobes, by comparison, appear to use a different set of genetic instructions in their development.
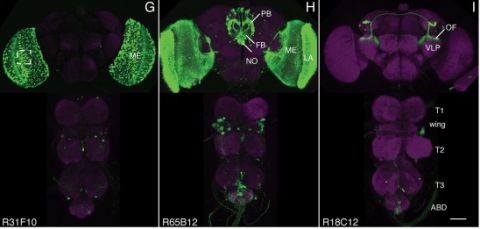
We established a collection of 7,000 transgenic lines of Drosophila melanogaster. Expression of GAL4 in each line is controlled by a different, defined fragment of genomic DNA that serves as a transcriptional enhancer. We used confocal microscopy of dissected nervous systems to determine the expression patterns driven by each fragment in the adult brain and ventral nerve cord. We present image data on 6,650 lines. Using both manual and machine-assisted annotation, we describe the expression patterns in the most useful lines. We illustrate the utility of these data for identifying novel neuronal cell types, revealing brain asymmetry, and describing the nature and extent of neuronal shape stereotypy. The GAL4 lines allow expression of exogenous genes in distinct, small subsets of the adult nervous system. The set of DNA fragments, each driving a documented expression pattern, will facilitate the generation of additional constructs for manipulating neuronal function. synapse was substantially elevated, at the endocytic zone there was no enhanced polymerization activity. We conclude that actin subserves spatially diverse, independently regulated processes throughout spines. Perisynaptic actin forms a uniquely dynamic structure well suited for direct, active regulation of the synapse.
For the overall strategy and methods used to produce the GAL4 lines:
Pfeiffer, B.D., Jenett, A., Hammonds, A.S., Ngo, T.T., Misra, S., Murphy, C., Scully, A., Carlson, J.W., Wan, K.H., Laverty, T.R., Mungall, C., Svirskas, R., Kadonaga, J.T., Doe, C.Q., Eisen, M.B., Celniker, S.E., Rubin, G.M. (2008). Tools for neuroanatomy and neurogenetics in Drosophila. Proc. Natl. Acad. Sci. USA 105, 9715-9720. http://www.pnas.org/content/105/28/9715.full.pdf+html synapse was substantially elevated, at the endocytic zone there was no enhanced polymerization activity. We conclude that actin subserves spatially diverse, independently regulated processes throughout spines. Perisynaptic actin forms a uniquely dynamic structure well suited for direct, active regulation of the synapse.
For data on expression in the embryo:
Manning, L., Purice, M.D., Roberts, J., Pollard, J.L., Bennett, A.L., Kroll, J.R., Dyukareva, A.V., Doan, P.N., Lupton, J.R., Strader, M.E., Tanner, S., Bauer, D., Wilbur, A., Tran, K.D., Laverty, T.R., Pearson, J.C., Crews, S.T., Rubin, G.M., and Doe, C.Q. (2012) Annotated embryonic CNS expression patterns of 5000 GMR GAL4 lines: a resource for manipulating gene expression and analyzing cis-regulatory motifs. Cell Reports (2012) Doi: 10.1016/j.celrep.2012.09.009 http://www.cell.com/cell-reports/fulltext/S2211-1247(12)00290-2 synapse was substantially elevated, at the endocytic zone there was no enhanced polymerization activity. We conclude that actin subserves spatially diverse, independently regulated processes throughout spines. Perisynaptic actin forms a uniquely dynamic structure well suited for direct, active regulation of the synapse.
For data on expression in imaginal discs:
Jory, A., Estella, C., Giorgianni, M.W., Slattery, M., Laverty, T.R., Rubin, G.M., and Mann, R.S. (2012) A survey of 6300 genomic fragments for cis-regulatory activity in the imaginal discs of Drosophila melanogaster. Cell Reports (2012) Doi: 10.1016/j.celrep.2012.09.010 http://www.cell.com/cell-reports/fulltext/S2211-1247(12)00291-4 synapse was substantially elevated, at the endocytic zone there was no enhanced polymerization activity. We conclude that actin subserves spatially diverse, independently regulated processes throughout spines. Perisynaptic actin forms a uniquely dynamic structure well suited for direct, active regulation of the synapse.
For data on expression in the larval nervous system:
Li, H.-H., Kroll, J.R., Lennox, S., Ogundeyi, O., Jeter, J., Depasquale, G., and Truman, J.W. (2013) A GAL4 driver resource for developmental and behavioral studies on the larval CNS of Drosophila. Cell Reports (submitted).
Fruit flies recognize hundreds of ecologically relevant odors and respond appropriately to them. The complexity, redundancy and interconnectedness of the olfactory machinery complicate efforts to pinpoint the functional contributions of any component neuron or receptor to behavior. Some contributions can only be elucidated in flies that carry multiple mutations and transgenes, but the production of such flies is currently labor-intensive and time-consuming. Here, we describe a set of transgenic flies that express the GAL80 in specific olfactory sensory neurons (). The GAL80s effectively and specifically subtract the activities of GAL4-driven transgenes that impart anatomical and physiological phenotypes. can allow researchers to efficiently activate only one or a few types of functional neurons in an otherwise nonfunctional olfactory background. Such experiments will improve our understanding of the mechanistic connections between odorant inputs and behavioral outputs at the resolution of only a few functional neurons.
We engineered electrochromic fluorescence resonance energy transfer (eFRET) genetically encoded voltage indicators (GEVIs) with “positive-going” fluorescence response to membrane depolarization through rational manipulation of the native proton transport pathway in microbial rhodopsins. We transformed the state-of-the-art eFRET GEVI Voltron into Positron, with kinetics and sensitivity equivalent to Voltron but flipped fluorescence signal polarity. We further applied this general approach to GEVIs containing different voltage sensitive rhodopsin domains and various fluorescent dye and fluorescent protein reporters.

Pushing the frontier of fluorescence microscopy requires the design of enhanced fluorophores with finely tuned properties. We recently discovered that incorporation of four-membered azetidine rings into classic fluorophore structures elicits substantial increases in brightness and photostability, resulting in the Janelia Fluor (JF) series of dyes. We refined and extended this strategy, finding that incorporation of 3-substituted azetidine groups allows rational tuning of the spectral and chemical properties of rhodamine dyes with unprecedented precision. This strategy allowed us to establish principles for fine-tuning the properties of fluorophores and to develop a palette of new fluorescent and fluorogenic labels with excitation ranging from blue to the far-red. Our results demonstrate the versatility of these new dyes in cells, tissues and animals.
Specific labeling of biomolecules with bright fluorophores is the keystone of fluorescence microscopy. Genetically encoded self-labeling tag proteins can be coupled to synthetic dyes inside living cells, resulting in brighter reporters than fluorescent proteins. Intracellular labeling using these techniques requires cell-permeable fluorescent ligands, however, limiting utility to a small number of classic fluorophores. Here we describe a simple structural modification that improves the brightness and photostability of dyes while preserving spectral properties and cell permeability. Inspired by molecular modeling, we replaced the N,N-dimethylamino substituents in tetramethylrhodamine with four-membered azetidine rings. This addition of two carbon atoms doubles the quantum efficiency and improves the photon yield of the dye in applications ranging from in vitro single-molecule measurements to super-resolution imaging. The novel substitution is generalizable, yielding a palette of chemical dyes with improved quantum efficiencies that spans the UV and visible range.
Fluorescence microscopy relies on dyes that absorb and then emit photons. In addition to fluorescence, fluorophores can undergo photochemical processes that decrease quantum yield or result in spectral shifts and irreversible photobleaching. Chemical strategies that suppress these undesirable pathways—thereby increasing the brightness and photostability of fluorophores—are crucial for advancing the frontier of bioimaging. Here, we describe a general method to improve small-molecule fluorophores by incorporating deuterium into the alkylamino auxochromes of rhodamines and other dyes. This strategy increases fluorescence quantum yield, inhibits photochemically induced spectral shifts, and slows irreparable photobleaching, yielding next-generation labels with improved performance in cellular imaging experiments.
Expanding the palette of fluorescent dyes is vital to push the frontier of biological imaging. Although rhodamine dyes remain the premier type of small-molecule fluorophore owing to their bioavailability and brightness, variants excited with far-red or near-infrared light suffer from poor performance due to their propensity to adopt a lipophilic, nonfluorescent form. We report a framework for rationalizing rhodamine behavior in biological environments and a general chemical modification for rhodamines that optimizes long-wavelength variants and enables facile functionalization with different chemical groups. This strategy yields red-shifted 'Janelia Fluor' (JF) dyes useful for biological imaging experiments in cells and in vivo.
Prolonged periods of forced social isolation is detrimental to well-being, yet we know little about which genes regulate susceptibility to its effects. In the fruit fly, Drosophila melanogaster, social isolation induces stark changes in behavior including increased aggression, locomotor activity, and resistance to ethanol sedation. To identify genes regulating sensitivity to isolation, I screened a collection of sixteen hundred P-element insertion lines for mutants with abnormal levels of all three isolation-induced behaviors. The screen identified three mutants whose affected genes are likely central to regulating the effects of isolation in flies. One mutant, sex pistol (sxp), became extremely aggressive and resistant to ethanol sedation when socially isolated. sxp also had a high level of male-male courtship. The mutation in sxp reduced the expression of two minor isoforms of the actin regulator hts (adducin), as well as mildly reducing expression of CalpA, a calcium-dependent protease. As a consequence, sxp also had increased expression of the insulin-like peptide, dILP5. Analysis of the social behavior of sxp suggests that these minor hts isoforms function to limit isolation-induced aggression, while chronically high levels of dILP5 increase male-male courtship.
The anatomy of many neural circuits is being characterized with increasing resolution, but their molecular properties remain mostly unknown. Here, we characterize gene expression patterns in distinct neural cell types of the visual system using genetic lines to access individual cell types, the TAPIN-seq method to measure their transcriptomes, and a probabilistic method to interpret these measurements. We used these tools to build a resource of high-resolution transcriptomes for 100 driver lines covering 67 cell types, available at http://www.opticlobe.com. Combining these transcriptomes with recently reported connectomes helps characterize how information is transmitted and processed across a range of scales, from individual synapses to circuit pathways. We describe examples that include identifying neurotransmitters, including cases of apparent co-release, generating functional hypotheses based on receptor expression, as well as identifying strong commonalities between different cell types.