Filter
Associated Lab
- Chklovskii Lab (3) Apply Chklovskii Lab filter
- Gonen Lab (1) Apply Gonen Lab filter
- Heberlein Lab (5) Apply Heberlein Lab filter
- Rubin Lab (2) Apply Rubin Lab filter
- Simpson Lab (2) Apply Simpson Lab filter
- Spruston Lab (2) Apply Spruston Lab filter
- Stern Lab (4) Apply Stern Lab filter
- Truman Lab (2) Apply Truman Lab filter
- Zuker Lab (3) Apply Zuker Lab filter
Publication Date
- December 2000 (3) Apply December 2000 filter
- November 2000 (3) Apply November 2000 filter
- October 2000 (3) Apply October 2000 filter
- August 2000 (3) Apply August 2000 filter
- June 2000 (4) Apply June 2000 filter
- April 2000 (3) Apply April 2000 filter
- March 2000 (6) Apply March 2000 filter
- February 2000 (1) Apply February 2000 filter
- January 2000 (3) Apply January 2000 filter
- Remove 2000 filter 2000
Type of Publication
29 Publications
Showing 21-29 of 29 resultsElectrophysiology and optical indicators have been used in vertebrate systems to investigate excitable cell firing and calcium transients, but both techniques have been difficult to apply in organisms with powerful reverse genetics. To overcome this limitation, we expressed cameleon proteins, genetically encoded calcium indicators, in the pharyngeal muscle of the nematode worm Caenorhabditis elegans. In intact transgenic animals expressing cameleons, fluorescence ratio changes accompanied muscular contraction, verifying detection of calcium transients. By comparing the magnitude and duration of calcium influx in wild-type and mutant animals, we were able to determine the effects of calcium channel proteins on pharyngeal calcium transients. We also successfully used cameleons to detect electrically evoked calcium transients in individual C. elegans neurons. This technique therefore should have broad applications in analyzing the regulation of excitable cell activity in genetically tractable organisms.
I consider a topographic projection between two neuronal layers with different densities of neurons. Given the number of output neurons connected to each input neuron (divergence) and the number of input neurons synapsing on each output neuron (convergence), I determine the widths of axonal and dendritic arbors which minimize the total volume of axons and dendrites. Analytical results for one-dimensional and two-dimensional projections can be summarized qualitatively in the following rule: neurons of the sparser layer should have arbors wider than those of the denser layer. This agrees with the anatomic data for retinal, cerebellar, olfactory bulb, and neocortical neurons the morphology and connectivity of which are known. The rule may be used to infer connectivity of neurons from their morphology.
Action potentials are the end product of synaptic integration, a process influenced by resting and active neuronal membrane properties. Diversity in these properties contributes to specialized mechanisms of synaptic integration and action potential firing, which are likely to be of functional significance within neural circuits. In the hippocampus, the majority of subicular pyramidal neurons fire high-frequency bursts of action potentials, whereas CA1 pyramidal neurons exhibit regular spiking behavior when subjected to direct somatic current injection. Using patch-clamp recordings from morphologically identified neurons in hippocampal slices, we analyzed and compared the resting and active membrane properties of pyramidal neurons in the subiculum and CA1 regions of the hippocampus. In response to direct somatic current injection, three subicular firing types were identified (regular spiking, weak bursting, and strong bursting), while all CA1 neurons were regular spiking. Within subiculum strong bursting neurons were found preferentially further away from the CA1 subregion. Input resistance (R(N)), membrane time constant (tau(m)), and depolarizing "sag" in response to hyperpolarizing current pulses were similar in all subicular neurons, while R(N) and tau(m) were significantly larger in CA1 neurons. The first spike of all subicular neurons exhibited similar action potential properties; CA1 action potentials exhibited faster rising rates, greater amplitudes, and wider half-widths than subicular action potentials. Therefore both the resting and active properties of CA1 pyramidal neurons are distinct from those of subicular neurons, which form a related class of neurons, differing in their propensity to burst. We also found that both regular spiking subicular and CA1 neurons could be transformed into a burst firing mode by application of a low concentration of 4-aminopyridine, suggesting that in both hippocampal subfields, firing properties are regulated by a slowly inactivating, D-type potassium current. The ability of all subicular pyramidal neurons to burst strengthens the notion that they form a single neuronal class, sharing a burst generating mechanism that is stronger in some cells than others.
Previous studies showed that Roundabout (Robo) in Drosophila is a repulsive axon guidance receptor that binds to Slit, a repellent secreted by midline glia. In robo mutants, growth cones cross and recross the midline, while, in slit mutants, growth cones enter the midline but fail to leave it. This difference suggests that Slit must have more than one receptor controlling midline guidance. In the absence of Robo, some other Slit receptor ensures that growth cones do not stay at the midline, even though they cross and recross it. Here we show that the Drosophila genome encodes three Robo receptors and that Robo and Robo2 have distinct functions, which together control repulsive axon guidance at the midline. The robo,robo2 double mutant is largely identical to slit.
Slit is secreted by midline glia in Drosophila and functions as a short-range repellent to control midline crossing. Although most Slit stays near the midline, some diffuses laterally, functioning as a long-range chemorepellent. Here we show that a combinatorial code of Robo receptors controls lateral position in the CNS by responding to this presumptive Slit gradient. Medial axons express only Robo, intermediate axons express Robo3 and Robo, while lateral axons express Robo2, Robo3, and Robo. Removal of robo2 or robo3 causes lateral axons to extend medially; ectopic expression of Robo2 or Robo3 on medial axons drives them laterally. Precise topography of longitudinal pathways appears to be controlled by a combination of long-range guidance (the Robo code determining region) and short-range guidance (discrete local cues determining specific location within a region).
Bitter taste perception provides animals with critical protection against ingestion of poisonous compounds. In the accompanying paper, we report the characterization of a large family of putative mammalian taste receptors (T2Rs). Here we use a heterologous expression system to show that specific T2Rs function as bitter taste receptors. A mouse T2R (mT2R-5) responds to the bitter tastant cycloheximide, and a human and a mouse receptor (hT2R-4 and mT2R-8) responded to denatonium and 6-n-propyl-2-thiouracil. Mice strains deficient in their ability to detect cycloheximide have amino acid substitutions in the mT2R-5 gene; these changes render the receptor significantly less responsive to cycloheximide. We also expressed mT2R-5 in insect cells and demonstrate specific tastant-dependent activation of gustducin, a G protein implicated in bitter signaling. Since a single taste receptor cell expresses a large repertoire of T2Rs, these findings provide a plausible explanation for the uniform bitter taste that is evoked by many structurally unrelated toxic compounds.
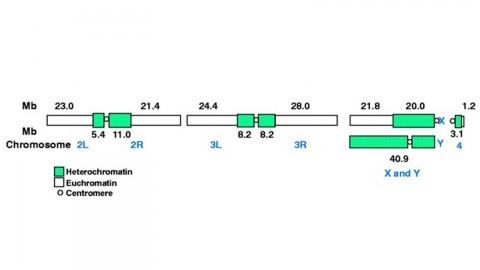
The fly Drosophila melanogaster is one of the most intensively studied organisms in biology and serves as a model system for the investigation of many developmental and cellular processes common to higher eukaryotes, including humans. We have determined the nucleotide sequence of nearly all of the approximately 120-megabase euchromatic portion of the Drosophila genome using a whole-genome shotgun sequencing strategy supported by extensive clone-based sequence and a high-quality bacterial artificial chromosome physical map. Efforts are under way to close the remaining gaps; however, the sequence is of sufficient accuracy and contiguity to be declared substantially complete and to support an initial analysis of genome structure and preliminary gene annotation and interpretation. The genome encodes approximately 13,600 genes, somewhat fewer than the smaller Caenorhabditis elegans genome, but with comparable functional diversity.
The steroid hormone 20-hydroxyecdysone (20E) initiates metamorphosis in insects by signaling through the ecdysone receptor complex, a heterodimer of the ecdysone receptor (EcR) and ultraspiracle (USP). Analysis of usp mutant clones in the wing disc of Drosophila shows that in the absence of USP, early hormone responsive genes such as EcR, DHR3 and E75B fail to up-regulate in response to 20E, but other genes that are normally expressed later, such as (&bgr;)-Ftz-F1 and the Z1 isoform of the Broad-Complex (BRC-Z1), are expressed precociously. Sensory neuron formation and axonal outgrowth, two early metamorphic events, also occur prematurely. In vitro experiments with cultured wing discs showed that BRC-Z1 expression and early metamorphic development are rendered steroid-independent in the usp mutant clones. These results are consistent with a model in which these latter processes are induced by a signal arising during the middle of the last larval stage but suppressed by the unliganded EcR/USP complex. Our observations suggest that silencing by the unliganded EcR/USP receptor and the subsequent release of silencing by moderate steroid levels may play an important role in coordinating early phases of steroid driven development.