Filter
Associated Lab
- Remove Rubin Lab filter Rubin Lab
Publication Date
Type of Publication
4 Publications
Showing 1-4 of 4 resultsFlexible goal-driven orientation requires that the position of a target be stored, especially in case the target moves out of sight. The capability to retain, recall and integrate such positional information into guiding behaviour has been summarized under the term spatial working memory. This kind of memory contains specific details of the presence that are not necessarily part of a long-term memory. Neurophysiological studies in primates indicate that sustained activity of neurons encodes the sensory information even though the object is no longer present. Furthermore they suggest that dopamine transmits the respective input to the prefrontal cortex, and simultaneous suppression by GABA spatially restricts this neuronal activity. Here we show that Drosophila melanogaster possesses a similar spatial memory during locomotion. Using a new detour setup, we show that flies can remember the position of an object for several seconds after it has been removed from their environment. In this setup, flies are temporarily lured away from the direction towards their hidden target, yet they are thereafter able to aim for their former target. Furthermore, we find that the GABAergic (stainable with antibodies against GABA) ring neurons of the ellipsoid body in the central brain are necessary and their plasticity is sufficient for a functional spatial orientation memory in flies. We also find that the protein kinase S6KII (ignorant) is required in a distinct subset of ring neurons to display this memory. Conditional expression of S6KII in these neurons only in adults can restore the loss of the orientation memory of the ignorant mutant. The S6KII signalling pathway therefore seems to be acutely required in the ring neurons for spatial orientation memory in flies.
Several aspects of locomotor control have been ascribed to the central complex of the insect brain; however, the role of distinct substructures of this complex is not well known. The tay bridge1 (tay1) mutant of Drosophila melanogaster was originally isolated on the basis of reduced walking speed and activity. In addition, tay1 is defective in the compensation of rotatory stimuli during walking and histologically, tay1 causes a mid-sagittal constriction of the protocerebral bridge, a constituent of the central complex. Cloning of the tay gene revealed that it encodes a novel protein with no significant homology to any known protein. To associate the behavioral phenotypes with the anatomical defect in the protocerebral bridge, we used different driver lines to express the tay cDNA in various neuronal subpopulations of the central brain in tay1-mutant flies. These experiments showed an association of the aberrant walking speed and activity with the structural defect in the protocerebral bridge. In contrast, the compensation of rotatory stimuli during walking was rescued without a restoration of the protocerebral bridge. The results of our differential rescue approach are supported by neuronal silencing experiments using conditional tetanus toxin expression in the same subset of neurons. These findings show for the first time that the walking speed and activity is controlled by different substructures of the central brain than the compensatory locomotion for rotatory stimuli.
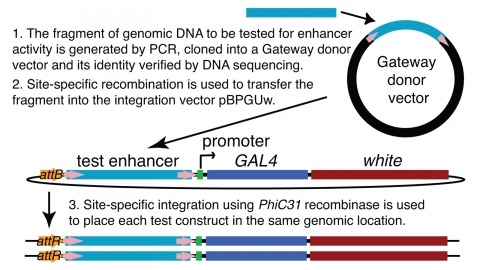
We demonstrate the feasibility of generating thousands of transgenic Drosophila melanogaster lines in which the expression of an exogenous gene is reproducibly directed to distinct small subsets of cells in the adult brain. We expect the expression patterns produced by the collection of 5,000 lines that we are currently generating to encompass all neurons in the brain in a variety of intersecting patterns. Overlapping 3-kb DNA fragments from the flanking noncoding and intronic regions of genes thought to have patterned expression in the adult brain were inserted into a defined genomic location by site-specific recombination. These fragments were then assayed for their ability to function as transcriptional enhancers in conjunction with a synthetic core promoter designed to work with a wide variety of enhancer types. An analysis of 44 fragments from four genes found that >80% drive expression patterns in the brain; the observed patterns were, on average, comprised of <100 cells. Our results suggest that the D. melanogaster genome contains >50,000 enhancers and that multiple enhancers drive distinct subsets of expression of a gene in each tissue and developmental stage. We expect that these lines will be valuable tools for neuroanatomy as well as for the elucidation of neuronal circuits and information flow in the fly brain.
The conditional expression of hairpin constructs in Drosophila melanogaster has emerged in recent years as a method of choice in functional genomic studies. To date, upstream activating site-driven RNA interference constructs have been inserted into the genome randomly using P-element-mediated transformation, which can result in false negatives due to variable expression. To avoid this problem, we have developed a transgenic RNA interference vector based on the phiC31 site-specific integration method.