Filter
Associated Lab
- Baker Lab (1) Apply Baker Lab filter
- Betzig Lab (2) Apply Betzig Lab filter
- Bock Lab (1) Apply Bock Lab filter
- Branson Lab (1) Apply Branson Lab filter
- Card Lab (2) Apply Card Lab filter
- Cardona Lab (3) Apply Cardona Lab filter
- Chklovskii Lab (4) Apply Chklovskii Lab filter
- Dickson Lab (1) Apply Dickson Lab filter
- Dudman Lab (1) Apply Dudman Lab filter
- Eddy/Rivas Lab (6) Apply Eddy/Rivas Lab filter
- Fetter Lab (5) Apply Fetter Lab filter
- Fitzgerald Lab (1) Apply Fitzgerald Lab filter
- Gonen Lab (1) Apply Gonen Lab filter
- Grigorieff Lab (3) Apply Grigorieff Lab filter
- Harris Lab (1) Apply Harris Lab filter
- Heberlein Lab (8) Apply Heberlein Lab filter
- Hess Lab (2) Apply Hess Lab filter
- Jayaraman Lab (2) Apply Jayaraman Lab filter
- Kainmueller Lab (6) Apply Kainmueller Lab filter
- Keller Lab (1) Apply Keller Lab filter
- Lavis Lab (2) Apply Lavis Lab filter
- Lee (Albert) Lab (1) Apply Lee (Albert) Lab filter
- Leonardo Lab (1) Apply Leonardo Lab filter
- Liu (Zhe) Lab (1) Apply Liu (Zhe) Lab filter
- Looger Lab (7) Apply Looger Lab filter
- Magee Lab (3) Apply Magee Lab filter
- Menon Lab (3) Apply Menon Lab filter
- Pastalkova Lab (2) Apply Pastalkova Lab filter
- Pavlopoulos Lab (2) Apply Pavlopoulos Lab filter
- Reiser Lab (1) Apply Reiser Lab filter
- Riddiford Lab (8) Apply Riddiford Lab filter
- Rubin Lab (2) Apply Rubin Lab filter
- Saalfeld Lab (4) Apply Saalfeld Lab filter
- Satou Lab (1) Apply Satou Lab filter
- Schreiter Lab (3) Apply Schreiter Lab filter
- Shroff Lab (2) Apply Shroff Lab filter
- Simpson Lab (1) Apply Simpson Lab filter
- Singer Lab (1) Apply Singer Lab filter
- Spruston Lab (4) Apply Spruston Lab filter
- Stern Lab (5) Apply Stern Lab filter
- Sternson Lab (2) Apply Sternson Lab filter
- Svoboda Lab (9) Apply Svoboda Lab filter
- Tjian Lab (4) Apply Tjian Lab filter
- Truman Lab (4) Apply Truman Lab filter
- Turaga Lab (1) Apply Turaga Lab filter
- Turner Lab (1) Apply Turner Lab filter
- Zlatic Lab (1) Apply Zlatic Lab filter
- Zuker Lab (2) Apply Zuker Lab filter
Associated Project Team
Publication Date
- December 2009 (14) Apply December 2009 filter
- November 2009 (4) Apply November 2009 filter
- October 2009 (17) Apply October 2009 filter
- September 2009 (9) Apply September 2009 filter
- August 2009 (14) Apply August 2009 filter
- July 2009 (16) Apply July 2009 filter
- June 2009 (16) Apply June 2009 filter
- May 2009 (10) Apply May 2009 filter
- April 2009 (11) Apply April 2009 filter
- March 2009 (11) Apply March 2009 filter
- February 2009 (13) Apply February 2009 filter
- January 2009 (23) Apply January 2009 filter
- Remove 2009 filter 2009
Type of Publication
158 Publications
Showing 11-20 of 158 resultsAfter spinal cord injury (SCI), the formation of glial scar contributes to the failure of injured adult axons to regenerate past the lesion. Increasing evidence indicates that olfactory ensheathing cells (OECs) implanted into spinal cord are found to migrate into the lesion site and induce axons regeneration beyond glial scar and resumption of functions. However, little is known about the mechanisms of OECs migrating from injection site to glial scar/lesion site.
Artificial lipidic bilayers are widely used as a model for the lipid matrix in biological cell membranes. We use the Pockels electro-optical effect to investigate the properties of an artificial lipidic membrane doped with nonlinear molecules in the outer layer. We report here what is believed to be the first electro-optical Pockels signal and image from such a membrane. The electro-optical dephasing distribution within the membrane is imaged and the signal is shown to be linear as a function of the applied voltage. A theoretical analysis taking into account the statistical orientation distribution of the inserted dye molecules allows us to estimate the doped membrane nonlinearity. Ongoing extensions of this work to living cell membranes are discussed.
The excitability of individual dendritic branches is a plastic property of neurons. We found that experience in an enriched environment increased propagation of dendritic Na(+) spikes in a subset of individual dendritic branches in rat hippocampal CA1 pyramidal neurons and that this effect was mainly mediated by localized downregulation of A-type K(+) channel function. Thus, dendritic plasticity might be used to store recent experience in individual branches of the dendritic arbor.
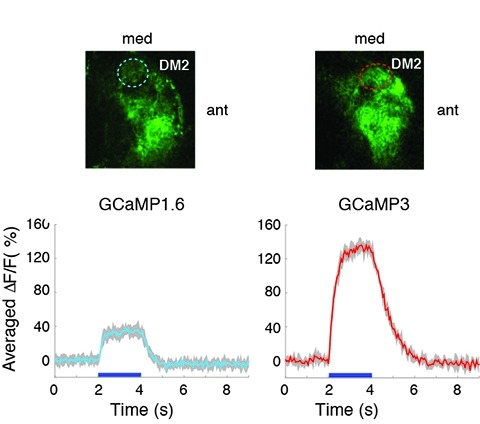
Genetically encoded calcium indicators (GECIs) can be used to image activity in defined neuronal populations. However, current GECIs produce inferior signals compared to synthetic indicators and recording electrodes, precluding detection of low firing rates. We developed a single-wavelength GCaMP2-based GECI (GCaMP3), with increased baseline fluorescence (3-fold), increased dynamic range (3-fold) and higher affinity for calcium (1.3-fold). We detected GCaMP3 fluorescence changes triggered by single action potentials in pyramidal cell dendrites, with signal-to-noise ratio and photostability substantially better than those of GCaMP2, D3cpVenus and TN-XXL. In Caenorhabditis elegans chemosensory neurons and the Drosophila melanogaster antennal lobe, sensory stimulation-evoked fluorescence responses were significantly enhanced with GCaMP3 (4-6-fold). In somatosensory and motor cortical neurons in the intact mouse, GCaMP3 detected calcium transients with amplitudes linearly dependent on action potential number. Long-term imaging in the motor cortex of behaving mice revealed large fluorescence changes in imaged neurons over months.
During the development of the central nervous system (CNS) of Drosophila, neuronal stem cells, the neuroblasts (NBs), first generate a set of highly diverse neurons, the primary neurons that mature to control larval behavior, and then more homogeneous sets of neurons that show delayed maturation and are primarily used in the adult. These latter, ’secondary’ neurons show a complex pattern of expression of broad, which encodes a transcription factor usually associated with metamorphosis, where it acts as a key regulator in the transitions from larva and pupa.
The dynamic evolution of organelle compartmentalization in eukaryotes and how strictly compartmentalization is maintained are matters of ongoing debate. While the endoplasmic reticulum (ER) is classically envisioned as the site of protein cotranslational translocation, it has recently been proposed to have pluripotent functions. Using transfected reporter constructs, organelle-specific markers, and functional enzyme assays, we now show that in an early-diverging protozoan, Giardia lamblia, endocytosis and subsequent degradation of exogenous proteins occur in the ER or in an adjacent and communicating compartment. The Giardia endomembrane system is simple compared to those of typical eukaryotes. It lacks peroxisomes, a classical Golgi apparatus, and canonical lysosomes. Giardia orthologues of mammalian lysosomal proteases function within an ER-like tubulovesicular compartment, which itself can dynamically communicate with clathrin-containing vacuoles at the periphery of the cell to receive endocytosed proteins. These primitive characteristics support Giardia's proposed early branching and could serve as a model to study the compartmentalization of endocytic and lysosomal functions into organelles distinct from the ER. This system also may have functional similarity to the retrograde transport of toxins and major histocompatibility complex class I function in the ER of mammals.
DNA sequencing-by-synthesis (SBS) technology, using a polymerase or ligase enzyme as its core biochemistry, has already been incorporated in several second-generation DNA sequencing systems with significant performance. Notwithstanding the substantial success of these SBS platforms, challenges continue to limit the ability to reduce the cost of sequencing a human genome to $100,000 or less. Achieving dramatically reduced cost with enhanced throughput and quality will require the seamless integration of scientific and technological effort across disciplines within biochemistry, chemistry, physics and engineering. The challenges include sample preparation, surface chemistry, fluorescent labels, optimizing the enzyme-substrate system, optics, instrumentation, understanding tradeoffs of throughput versus accuracy, and read-length/phasing limitations. By framing these challenges in a manner accessible to a broad community of scientists and engineers, we hope to solicit input from the broader research community on means of accelerating the advancement of genome sequencing technology.
Wnt signaling through Frizzled proteins guides posterior cells and axons in C. elegans into different spatial domains. Here we demonstrate an essential role for Wnt signaling through Ror tyrosine kinase homologs in the most prominent anterior neuropil, the nerve ring. A genetic screen uncovered cwn-2, the C. elegans homolog of Wnt5, as a regulator of nerve ring placement. In cwn-2 mutants, all neuronal structures in and around the nerve ring are shifted to an abnormal anterior position. cwn-2 is required at the time of nerve ring formation; it is expressed by cells posterior of the nerve ring, but its precise site of expression is not critical for its function. In nerve ring development, cwn-2 acts primarily through the Wnt receptor CAM-1 (Ror), together with the Frizzled protein MIG-1, with parallel roles for the Frizzled protein CFZ-2. The identification of CAM-1 as a CWN-2 receptor contrasts with CAM-1 action as a non-receptor in other C. elegans Wnt pathways. Cell-specific rescue of cam-1 and cell ablation experiments reveal a crucial role for the SIA and SIB neurons in positioning the nerve ring, linking Wnt signaling to specific cells that organize the anterior nervous system.
The C. elegans cell lineage provides a unique opportunity to look at how cell lineage affects patterns of gene expression. We developed an automatic cell lineage analyzer that converts high-resolution images of worms into a data table showing fluorescence expression with single-cell resolution. We generated expression profiles of 93 genes in 363 specific cells from L1 stage larvae and found that cells with identical fates can be formed by different gene regulatory pathways. Molecular signatures identified repeating cell fate modules within the cell lineage and enabled the generation of a molecular differentiation map that reveals points in the cell lineage when developmental fates of daughter cells begin to diverge. These results demonstrate insights that become possible using computational approaches to analyze quantitative expression from many genes in parallel using a digital gene expression atlas.
Forkhead transcription factors play critical roles in leukocyte homeostasis. To study further the immunological functions of Foxo1, we generated mice that selectively lack Foxo1 in T cells (Foxo1(flox/flox) Lck.cre(+)conditional knockout mice (cKO)). Although thymocyte development appeared relatively normal, Foxo1 cKO mice harbored significantly increased percentages of mature single positive T cells in the thymus as compared with WT mice, yet possessed smaller lymph nodes and spleens that contained fewer T cells. Foxo1 cKO T cells were not more prone to apoptosis, but instead were characterized by a CD62L(lo) CCR7(lo) CD44(hi) surface phenotype, a poorly populated lymphoid compartment in the periphery, and were relatively refractory to TCR stimulation, all of which were associated with reduced expression of Sell, Klf2, Ccr7, and S1pr1. Thus, Foxo1 is critical for naïve T cells to populate the peripheral lymphoid organs by coordinating a molecular program that maintains homeostasis and regulates trafficking.