Filter
Associated Lab
- Ahrens Lab (4) Apply Ahrens Lab filter
- Betzig Lab (2) Apply Betzig Lab filter
- Branson Lab (1) Apply Branson Lab filter
- Darshan Lab (3) Apply Darshan Lab filter
- Druckmann Lab (5) Apply Druckmann Lab filter
- Dudman Lab (3) Apply Dudman Lab filter
- Fetter Lab (1) Apply Fetter Lab filter
- Freeman Lab (3) Apply Freeman Lab filter
- Harris Lab (6) Apply Harris Lab filter
- Hermundstad Lab (1) Apply Hermundstad Lab filter
- Jayaraman Lab (9) Apply Jayaraman Lab filter
- Ji Lab (2) Apply Ji Lab filter
- Karpova Lab (2) Apply Karpova Lab filter
- Lavis Lab (4) Apply Lavis Lab filter
- Lee (Albert) Lab (3) Apply Lee (Albert) Lab filter
- Leonardo Lab (2) Apply Leonardo Lab filter
- Liu (Zhe) Lab (1) Apply Liu (Zhe) Lab filter
- Looger Lab (20) Apply Looger Lab filter
- Magee Lab (1) Apply Magee Lab filter
- Pachitariu Lab (2) Apply Pachitariu Lab filter
- Pastalkova Lab (1) Apply Pastalkova Lab filter
- Podgorski Lab (2) Apply Podgorski Lab filter
- Romani Lab (5) Apply Romani Lab filter
- Rubin Lab (3) Apply Rubin Lab filter
- Saalfeld Lab (2) Apply Saalfeld Lab filter
- Schreiter Lab (13) Apply Schreiter Lab filter
- Spruston Lab (3) Apply Spruston Lab filter
- Sternson Lab (4) Apply Sternson Lab filter
- Remove Svoboda Lab filter Svoboda Lab
- Tillberg Lab (3) Apply Tillberg Lab filter
- Turner Lab (3) Apply Turner Lab filter
Associated Project Team
Publication Date
- 2023 (5) Apply 2023 filter
- 2022 (6) Apply 2022 filter
- 2021 (7) Apply 2021 filter
- 2020 (5) Apply 2020 filter
- 2019 (14) Apply 2019 filter
- 2018 (11) Apply 2018 filter
- 2017 (9) Apply 2017 filter
- 2016 (8) Apply 2016 filter
- 2015 (9) Apply 2015 filter
- 2014 (7) Apply 2014 filter
- 2013 (10) Apply 2013 filter
- 2012 (9) Apply 2012 filter
- 2011 (7) Apply 2011 filter
- 2010 (7) Apply 2010 filter
- 2009 (9) Apply 2009 filter
- 2008 (6) Apply 2008 filter
- 2007 (3) Apply 2007 filter
- 2005 (2) Apply 2005 filter
- 2004 (1) Apply 2004 filter
Type of Publication
135 Publications
Showing 101-110 of 135 resultsIn the rodent vibrissal system, active sensation and sensorimotor integration are mediated in part by connections between barrel cortex and vibrissal motor cortex. Little is known about how these structures interact at the level of neurons. We used Channelrhodopsin-2 (ChR2) expression, combined with anterograde and retrograde labeling, to map connections between barrel cortex and pyramidal neurons in mouse motor cortex. Barrel cortex axons preferentially targeted upper layer (L2/3, L5A) neurons in motor cortex; input to neurons projecting back to barrel cortex was particularly strong. Barrel cortex input to deeper layers (L5B, L6) of motor cortex, including neurons projecting to the brainstem, was weak, despite pronounced geometric overlap of dendrites with axons from barrel cortex. Neurons in different layers received barrel cortex input within stereotyped dendritic domains. The cortico-cortical neurons in superficial layers of motor cortex thus couple motor and sensory signals and might mediate sensorimotor integration and motor learning.
Concomitant with the publication of this Special Issue of Neuroinformatics, a substantially updated version of the DIADEM web site has been released at http://diademchallenge.org. This web site was originally designed to host the challenge for automating the digital reconstruction of axonal and dendritic morphology (hence the DIADEM acronym). This post-competition version features additional content for continued use as the access point for DIADEM-related material. From the very beginning, one of the spirits of DIADEM has been to share data and resources with the neuroscience research community at large. The resources available from or linked to the DIADEM website constitute a substantial scientific legacy of the 2009/2010 competition. The new content includes finalist algorithms, image stack data, gold standard reconstructions, an updated DIADEM metric, and a retrospective on the competition in text and images.

Site-specific recombinases have been used for two decades to manipulate the structure of animal genomes in highly predictable ways and have become major research tools. However, the small number of recombinases demonstrated to have distinct specificities, low toxicity, and sufficient activity to drive reactions to completion in animals has been a limitation. In this report we show that four recombinases derived from yeast-KD, B2, B3, and R-are highly active and nontoxic in Drosophila and that KD, B2, B3, and the widely used FLP recombinase have distinct target specificities. We also show that the KD and B3 recombinases are active in mice.
Rodents move their whiskers to locate and identify objects. Cortical areas involved in vibrissal somatosensation and sensorimotor integration include the vibrissal area of the primary motor cortex (vM1), primary somatosensory cortex (vS1; barrel cortex), and secondary somatosensory cortex (S2). We mapped local excitatory pathways in each area across all cortical layers using glutamate uncaging and laser scanning photostimulation. We analyzed these maps to derive laminar connectivity matrices describing the average strengths of pathways between individual neurons in different layers and between entire cortical layers. In vM1, the strongest projection was L2/3→L5. In vS1, strong projections were L2/3→L5 and L4→L3. L6 input and output were weak in both areas. In S2, L2/3→L5 exceeded the strength of the ascending L4→L3 projection, and local input to L6 was prominent. The most conserved pathways were L2/3→L5, and the most variable were L4→L2/3 and pathways involving L6. Local excitatory circuits in different cortical areas are organized around a prominent descending pathway from L2/3→L5, suggesting that sensory cortices are elaborations on a basic motor cortex-like plan.
Optogenetics is routinely used to activate and inactivate genetically defined neuronal populations in vivo. A second optogenetic revolution will occur when spatially distributed and sparse neural assemblies can be precisely manipulated in behaving animals.
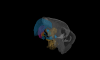
The basal ganglia play a critical role in the regulation of voluntary action in vertebrates. Our understanding of the function of the basal ganglia relies heavily upon anatomical information, but continued progress will require an understanding of the specific functional roles played by diverse cell types and their connectivity. An increasing number of mouse lines allow extensive identification, characterization, and manipulation of specified cell types in the basal ganglia. Despite the promise of genetically modified mice for elucidating the functional roles of diverse cell types, there is relatively little anatomical data obtained directly in the mouse. Here we have characterized the retrograde labeling obtained from a series of tracer injections throughout the dorsal striatum of adult mice. We found systematic variations in input along both the medial-lateral and anterior-posterior neuraxes in close agreement with canonical features of basal ganglia anatomy in the rat. In addition to the canonical features we have provided experimental support for the importance of non-canonical inputs to the striatum from the raphe nuclei and the amygdala. To look for organization at a finer scale we have analyzed the correlation structure of labeling intensity across our entire dataset. Using this analysis we found substantial local heterogeneity within the large-scale order. From this analysis we conclude that individual striatal sites receive varied combinations of cortical and thalamic input from multiple functional areas, consistent with some earlier studies in the rat that have suggested the presence of a combinatorial map.
Spatial navigation is often used as a behavioral task in studies of the neuronal circuits that underlie cognition, learning and memory in rodents. The combination of in vivo microscopy with genetically encoded indicators has provided an important new tool for studying neuronal circuits, but has been technically difficult to apply during navigation. Here we describe methods for imaging the activity of neurons in the CA1 region of the hippocampus with subcellular resolution in behaving mice. Neurons that expressed the genetically encoded calcium indicator GCaMP3 were imaged through a chronic hippocampal window. Head-restrained mice performed spatial behaviors in a setup combining a virtual reality system and a custom-built two-photon microscope. We optically identified populations of place cells and determined the correlation between the location of their place fields in the virtual environment and their anatomical location in the local circuit. The combination of virtual reality and high-resolution functional imaging should allow a new generation of studies to investigate neuronal circuit dynamics during behavior.
The primary auditory cortex (A1) is organized tonotopically, with neurons sensitive to high and low frequencies arranged in a rostro-caudal gradient. We used laser scanning photostimulation in acute slices to study the organization of local excitatory connections onto layers 2 and 3 (L2/3) of the mouse A1. Consistent with the organization of other cortical regions, synaptic inputs along the isofrequency axis (orthogonal to the tonotopic axis) arose predominantly within a column. By contrast, we found that local connections along the tonotopic axis differed from those along the isofrequency axis: some input pathways to L3 (but not L2) arose predominantly out-of-column. In vivo cell-attached recordings revealed differences between the sound-responsiveness of neurons in L2 and L3. Our results are consistent with the hypothesis that auditory cortical microcircuitry is specialized to the one-dimensional representation of frequency in the auditory cortex.
Classical studies have related the spiking of selected neocortical neurons to behavior, but little is known about activity sampled from the entire neural population. We recorded from neurons selected independent of spiking, using cell-attached recordings and two-photon calcium imaging, in the barrel cortex of mice performing an object localization task. Spike rates varied across neurons, from silence to >60 Hz. Responses were diverse, with some neurons showing large increases in spike rate when whiskers contacted the object. Nearly half the neurons discriminated object location; a small fraction of neurons discriminated perfectly. More active neurons were more discriminative. Layer (L) 4 and L5 contained the highest fractions of discriminating neurons (\~{}63% and 79%, respectively), but a few L2/3 neurons were also highly discriminating. Approximately 13,000 spikes per activated barrel column were available to mice for decision making. Coding of object location in the barrel cortex is therefore highly redundant.