Filter
Associated Lab
- Aguilera Castrejon Lab (1) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (53) Apply Ahrens Lab filter
- Aso Lab (40) Apply Aso Lab filter
- Baker Lab (19) Apply Baker Lab filter
- Betzig Lab (101) Apply Betzig Lab filter
- Beyene Lab (8) Apply Beyene Lab filter
- Bock Lab (14) Apply Bock Lab filter
- Branson Lab (50) Apply Branson Lab filter
- Card Lab (36) Apply Card Lab filter
- Cardona Lab (45) Apply Cardona Lab filter
- Chklovskii Lab (10) Apply Chklovskii Lab filter
- Clapham Lab (14) Apply Clapham Lab filter
- Cui Lab (19) Apply Cui Lab filter
- Darshan Lab (8) Apply Darshan Lab filter
- Dickson Lab (32) Apply Dickson Lab filter
- Druckmann Lab (21) Apply Druckmann Lab filter
- Dudman Lab (38) Apply Dudman Lab filter
- Eddy/Rivas Lab (30) Apply Eddy/Rivas Lab filter
- Egnor Lab (4) Apply Egnor Lab filter
- Espinosa Medina Lab (15) Apply Espinosa Medina Lab filter
- Feliciano Lab (7) Apply Feliciano Lab filter
- Fetter Lab (31) Apply Fetter Lab filter
- Fitzgerald Lab (16) Apply Fitzgerald Lab filter
- Freeman Lab (15) Apply Freeman Lab filter
- Funke Lab (38) Apply Funke Lab filter
- Gonen Lab (59) Apply Gonen Lab filter
- Grigorieff Lab (34) Apply Grigorieff Lab filter
- Harris Lab (53) Apply Harris Lab filter
- Heberlein Lab (13) Apply Heberlein Lab filter
- Hermundstad Lab (23) Apply Hermundstad Lab filter
- Hess Lab (74) Apply Hess Lab filter
- Ilanges Lab (2) Apply Ilanges Lab filter
- Jayaraman Lab (42) Apply Jayaraman Lab filter
- Ji Lab (33) Apply Ji Lab filter
- Johnson Lab (1) Apply Johnson Lab filter
- Karpova Lab (13) Apply Karpova Lab filter
- Keleman Lab (8) Apply Keleman Lab filter
- Keller Lab (61) Apply Keller Lab filter
- Koay Lab (2) Apply Koay Lab filter
- Lavis Lab (137) Apply Lavis Lab filter
- Lee (Albert) Lab (29) Apply Lee (Albert) Lab filter
- Leonardo Lab (19) Apply Leonardo Lab filter
- Li Lab (4) Apply Li Lab filter
- Lippincott-Schwartz Lab (97) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (1) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (58) Apply Liu (Zhe) Lab filter
- Looger Lab (137) Apply Looger Lab filter
- Magee Lab (31) Apply Magee Lab filter
- Menon Lab (12) Apply Menon Lab filter
- Murphy Lab (6) Apply Murphy Lab filter
- O'Shea Lab (6) Apply O'Shea Lab filter
- Otopalik Lab (1) Apply Otopalik Lab filter
- Pachitariu Lab (36) Apply Pachitariu Lab filter
- Pastalkova Lab (5) Apply Pastalkova Lab filter
- Pavlopoulos Lab (7) Apply Pavlopoulos Lab filter
- Pedram Lab (4) Apply Pedram Lab filter
- Podgorski Lab (16) Apply Podgorski Lab filter
- Reiser Lab (45) Apply Reiser Lab filter
- Riddiford Lab (20) Apply Riddiford Lab filter
- Romani Lab (31) Apply Romani Lab filter
- Rubin Lab (105) Apply Rubin Lab filter
- Saalfeld Lab (46) Apply Saalfeld Lab filter
- Satou Lab (1) Apply Satou Lab filter
- Scheffer Lab (36) Apply Scheffer Lab filter
- Schreiter Lab (51) Apply Schreiter Lab filter
- Sgro Lab (1) Apply Sgro Lab filter
- Shroff Lab (31) Apply Shroff Lab filter
- Simpson Lab (18) Apply Simpson Lab filter
- Singer Lab (37) Apply Singer Lab filter
- Spruston Lab (57) Apply Spruston Lab filter
- Stern Lab (73) Apply Stern Lab filter
- Sternson Lab (47) Apply Sternson Lab filter
- Stringer Lab (33) Apply Stringer Lab filter
- Svoboda Lab (131) Apply Svoboda Lab filter
- Tebo Lab (9) Apply Tebo Lab filter
- Tervo Lab (9) Apply Tervo Lab filter
- Tillberg Lab (18) Apply Tillberg Lab filter
- Tjian Lab (17) Apply Tjian Lab filter
- Truman Lab (58) Apply Truman Lab filter
- Turaga Lab (39) Apply Turaga Lab filter
- Turner Lab (27) Apply Turner Lab filter
- Vale Lab (7) Apply Vale Lab filter
- Voigts Lab (3) Apply Voigts Lab filter
- Wang (Meng) Lab (21) Apply Wang (Meng) Lab filter
- Wang (Shaohe) Lab (6) Apply Wang (Shaohe) Lab filter
- Wu Lab (8) Apply Wu Lab filter
- Zlatic Lab (26) Apply Zlatic Lab filter
- Zuker Lab (5) Apply Zuker Lab filter
Associated Project Team
- CellMap (12) Apply CellMap filter
- COSEM (3) Apply COSEM filter
- FIB-SEM Technology (3) Apply FIB-SEM Technology filter
- Fly Descending Interneuron (11) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (14) Apply Fly Functional Connectome filter
- Fly Olympiad (5) Apply Fly Olympiad filter
- FlyEM (53) Apply FlyEM filter
- FlyLight (49) Apply FlyLight filter
- GENIE (46) Apply GENIE filter
- Integrative Imaging (4) Apply Integrative Imaging filter
- Larval Olympiad (2) Apply Larval Olympiad filter
- MouseLight (18) Apply MouseLight filter
- NeuroSeq (1) Apply NeuroSeq filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (26) Apply Tool Translation Team (T3) filter
- Transcription Imaging (45) Apply Transcription Imaging filter
Publication Date
- 2025 (130) Apply 2025 filter
- 2024 (215) Apply 2024 filter
- 2023 (159) Apply 2023 filter
- 2022 (167) Apply 2022 filter
- 2021 (175) Apply 2021 filter
- 2020 (177) Apply 2020 filter
- 2019 (177) Apply 2019 filter
- 2018 (206) Apply 2018 filter
- 2017 (186) Apply 2017 filter
- 2016 (191) Apply 2016 filter
- 2015 (195) Apply 2015 filter
- 2014 (190) Apply 2014 filter
- 2013 (136) Apply 2013 filter
- 2012 (112) Apply 2012 filter
- 2011 (98) Apply 2011 filter
- 2010 (61) Apply 2010 filter
- 2009 (56) Apply 2009 filter
- 2008 (40) Apply 2008 filter
- 2007 (21) Apply 2007 filter
- 2006 (3) Apply 2006 filter
Type of Publication
- Remove Janelia filter Janelia
2695 Publications
Showing 2091-2100 of 2695 resultsElectron diffraction of extremely small three-dimensional crystals (MicroED) allows for structure determination from crystals orders of magnitude smaller than those used for X-ray crystallography. MicroED patterns, which are collected in a transmission electron microscope, were initially not amenable to indexing and intensity extraction by standard software, which necessitated the development of a suite of programs for data processing. The MicroED suite was developed to accomplish the tasks of unit-cell determination, indexing, background subtraction, intensity measurement and merging, resulting in data that can be carried forward to molecular replacement and structure determination. This ad hoc solution has been modified for more general use to provide a means for processing MicroED data until the technique can be fully implemented into existing crystallographic software packages. The suite is written in Python and the source code is available under a GNU General Public License.
The evolution of behavior seems inconsistent with the deep homology of neuromodulatory signaling. G protein coupled receptors (GPCRs) evolved slowly from a common ancestor through a process involving gene duplication, neofunctionalization, and loss. Neuropeptides co-evolved with their receptors and exhibit many conserved functions. Furthermore, brain areas are highly conserved with suggestions of deep anatomical homology between arthropods and vertebrates. Yet, behavior evolved more rapidly; even members of the same genus or species can differ in heritable behavior. The solution to the paradox involves changes in the compartmentalization, or subfunctionalization, of neuromodulation; neurons shift their expression of GPCRs and the content of monoamines and neuropeptides. Furthermore, parallel evolution of neuromodulatory signaling systems suggests a route for repeated evolution of similar behaviors.
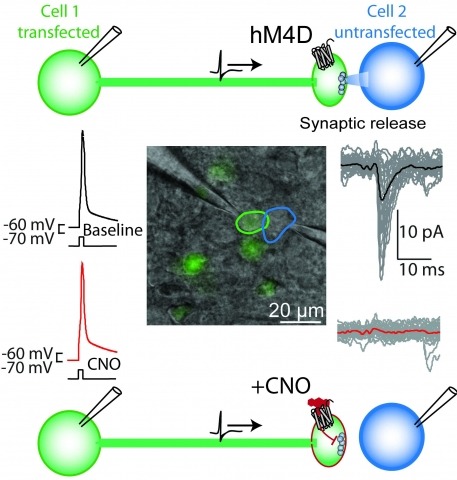
Brain function is mediated by neural circuit connectivity, and elucidating the role of connections is aided by techniques to block their output. We developed cell-type-selective, reversible synaptic inhibition tools for mammalian neural circuits by leveraging G protein signaling pathways to suppress synaptic vesicle release. Here, we find that the pharmacologically selective designer Gi-protein-coupled receptor hM4D is a presynaptic silencer in the presence of its cognate ligand clozapine-N-oxide (CNO). Activation of hM4D signaling sharply reduced synaptic release probability and synaptic current amplitude. To demonstrate the utility of this tool for neural circuit perturbations, we developed an axon-selective hM4D-neurexin variant and used spatially targeted intracranial CNO injections to localize circuit connections from the hypothalamus to the midbrain responsible for feeding behavior. This synaptic silencing approach is broadly applicable for cell-type-specific and axon projection-selective functional analysis of diverse neural circuits.
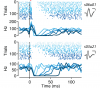
Dysfunction of the basal ganglia produces severe deficits in the timing, initiation, and vigor of movement. These diverse impairments suggest a control system gone awry. In engineered systems, feedback is critical for control. By contrast, models of the basal ganglia highlight feedforward circuitry and ignore intrinsic feedback circuits. In this study, we show that feedback via axon collaterals of substantia nigra projection neurons control the gain of the basal ganglia output. Through a combination of physiology, optogenetics, anatomy, and circuit mapping, we elaborate a general circuit mechanism for gain control in a microcircuit lacking interneurons. Our data suggest that diverse tonic firing rates, weak unitary connections and a spatially diffuse collateral circuit with distinct topography and kinetics from feedforward input is sufficient to implement divisive feedback inhibition. The importance of feedback for engineered systems implies that the intranigral microcircuit, despite its absence from canonical models, could be essential to basal ganglia function. DOI: http://dx.doi.org/10.7554/eLife.02397.001.

An important strategy for efficient neural coding is to match the range of cellular responses to the distribution of relevant input signals. However, the structure and relevance of sensory signals depend on behavioral state. Here, we show that behavior modifies neural activity at the earliest stages of fly vision. We describe a class of wide-field neurons that provide feedback to the most peripheral layer of the Drosophila visual system, the lamina. Using in vivo patch-clamp electrophysiology, we found that lamina wide-field neurons respond to low-frequency luminance fluctuations. Recordings in flying flies revealed that the gain and frequency tuning of wide-field neurons change during flight, and that these effects are mimicked by the neuromodulator octopamine. Genetically silencing wide-field neurons increased behavioral responses to slow-motion stimuli. Together, these findings identify a cell type that is gated by behavior to enhance neural coding by subtracting low-frequency signals from the inputs to motion detection circuits.
The budding yeast centromere contains Cse4, a specialized histone H3 variant. Fluorescence pulse-chase analysis of an internally tagged Cse4 reveals that it is replaced with newly synthesized molecules in S phase, remaining stably associated with centromeres thereafter. In contrast, C-terminally-tagged Cse4 is functionally impaired, showing slow cell growth, cell lethality at elevated temperatures, and extra-centromeric nuclear accumulation. Recent studies using such strains gave conflicting findings regarding the centromeric abundance and cell cycle dynamics of Cse4. Our findings indicate that internally tagged Cse4 is a better reporter of the biology of this histone variant. Furthermore, the size of centromeric Cse4 clusters was precisely mapped with a new 3D-PALM method, revealing substantial compaction during anaphase. Cse4-specific chaperone Scm3 displays steady-state, stoichiometric co-localization with Cse4 at centromeres throughout the cell cycle, while undergoing exchange with a nuclear pool. These findings suggest that a stable Cse4 nucleosome is maintained by dynamic chaperone-in-residence Scm3.DOI: http://dx.doi.org/10.7554/eLife.02203.001.
BACKGROUND: During courtship, male Drosophila melanogaster sing a multipart courtship song to female flies. This song is of particular interest because (1) it is species specific and varies widely within the genus, (2) it is a gating stimulus for females, who are sensitive detectors of conspecific song, and (3) it is the only sexual signal that is under both neural and genetic control. This song is perceived via mechanosensory neurons in the antennal Johnston's organ, which innervate the antennal mechanosensory and motor center (AMMC) of the brain. However, AMMC outputs that are responsible for detection and discrimination of conspecific courtship song remain unknown. RESULTS: Using a large-scale anatomical screen of AMMC interneurons, we identify seven projection neurons (aPNs) and five local interneurons (aLNs) that outline a complex architecture for the ascending mechanosensory pathway. Neuronal inactivation and hyperactivation during behavior reveal that only two classes of interneurons are necessary for song responses--the projection neuron aPN1 and GABAergic interneuron aLN(al). These neurons are necessary in both male and female flies. Physiological recordings in aPN1 reveal the integration of courtship song as a function of pulse rate and outline an intracellular transfer function that likely facilitates the response to conspecific song. CONCLUSIONS: These results reveal a critical pathway for courtship hearing in male and female flies, in which both aLN(al) and aPN1 mediate the detection of conspecific song. The pathways arising from these neurons likely serve as a critical neural substrate for behavioral reproductive isolation in D. melanogaster.
Nonmuscle myosin II (NM II) powers myriad developmental and cellular processes, including embryogenesis, cell migration, and cytokinesis [1]. To exert its functions, monomers of NM II assemble into bipolar filaments that produce a contractile force on the actin cytoskeleton. Mammalian cells express up to three isoforms of NM II (NM IIA, IIB, and IIC), each of which possesses distinct biophysical properties and supports unique as well as redundant cellular functions [2-8]. Despite previous efforts [9-13], it remains unclear whether NM II isoforms assemble in living cells to produce mixed (heterotypic) bipolar filaments or whether filaments consist entirely of a single isoform (homotypic). We addressed this question using fluorescently tagged versions of NM IIA, IIB, and IIC, isoform-specific immunostaining of the endogenous proteins, and two-color total internal reflection fluorescence structured-illumination microscopy, or TIRF-SIM, to visualize individual myosin II bipolar filaments inside cells. We show that NM II isoforms coassemble into heterotypic filaments in a variety of settings, including various types of stress fibers, individual filaments throughout the cell, and the contractile ring. We also show that the differential distribution of NM IIA and NM IIB typically seen in confocal micrographs of well-polarized cells is reflected in the composition of individual bipolar filaments. Interestingly, this differential distribution is less pronounced in freshly spread cells, arguing for the existence of a sorting mechanism acting over time. Together, our work argues that individual NM II isoforms are potentially performing both isoform-specific and isoform-redundant functions while coassembled with other NM II isoforms.
The role of juvenile hormone (JH) in regulating the timing and nature of insect molts is well-established. Increasing evidence suggests that JH is also involved in regulating final insect size. Here we elucidate the developmental mechanism through which JH regulates body size in developing Drosophila larvae by genetically ablating the JH-producing organ, the corpora allata (CA). We found that larvae that lack CA pupariated at smaller sizes than control larvae due to a reduced larval growth rate. Neither the timing of the metamorphic molt nor the duration of larval growth was affected by the loss of JH. Further, we show that the effects of JH on growth rate are dependent on the forkhead box O transcription factor (FOXO), which is negatively regulated by the insulin-signaling pathway. Larvae that lacked the CA had elevated levels of FOXO activity, whereas a loss-of-function mutation of FOXO rescued the effects of CA ablation on final body size. Finally, the effect of JH on growth appears to be mediated, at least in part, via ecdysone synthesis in the prothoracic gland. These results indicate a role of JH in regulating growth rate via the ecdysone- and insulin-signaling pathways.

Visual motion perception is critical to many animal behaviors, and flies have emerged as a powerful model system for exploring this fundamental neural computation. Although numerous studies have suggested that fly motion vision is governed by a simple neural circuit [1-3], the implementation of this circuit has remained mysterious for decades. Connectomics and neurogenetics have produced a surge in recent progress, and several studies have shown selectivity for light increments (ON) or decrements (OFF) in key elements associated with this circuit [4-7]. However, related studies have reached disparate conclusions about where this selectivity emerges and whether it plays a major role in motion vision [8-13]. To address these questions, we examined activity in the neuropil thought to be responsible for visual motion detection, the medulla, of Drosophila melanogaster in response to a range of visual stimuli using two-photon calcium imaging. We confirmed that the input neurons of the medulla, the LMCs, are not responsible for light-on and light-off selectivity. We then examined the pan-neural response of medulla neurons and found prominent selectivity for light-on and light-off in layers of the medulla associated with two anatomically derived pathways (L1/L2 associated) [14, 15]. We next examined the activity of prominent interneurons within each pathway (Mi1 and Tm1) and found that these neurons have corresponding selectivity for light-on or light-off. These results provide direct evidence that motion is computed in parallel light-on and light-off pathways, demonstrate that this selectivity emerges in neurons immediately downstream of the LMCs, and specify where crucial elements of motion computation occur.