Filter
Associated Lab
- Aguilera Castrejon Lab (15) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (11) Apply Ahrens Lab filter
- Baker Lab (19) Apply Baker Lab filter
- Betzig Lab (12) Apply Betzig Lab filter
- Beyene Lab (5) Apply Beyene Lab filter
- Bock Lab (3) Apply Bock Lab filter
- Branson Lab (3) Apply Branson Lab filter
- Card Lab (6) Apply Card Lab filter
- Cardona Lab (19) Apply Cardona Lab filter
- Chklovskii Lab (3) Apply Chklovskii Lab filter
- Clapham Lab (1) Apply Clapham Lab filter
- Darshan Lab (4) Apply Darshan Lab filter
- Dennis Lab (1) Apply Dennis Lab filter
- Dickson Lab (14) Apply Dickson Lab filter
- Druckmann Lab (4) Apply Druckmann Lab filter
- Dudman Lab (12) Apply Dudman Lab filter
- Egnor Lab (7) Apply Egnor Lab filter
- Espinosa Medina Lab (4) Apply Espinosa Medina Lab filter
- Fetter Lab (10) Apply Fetter Lab filter
- Fitzgerald Lab (13) Apply Fitzgerald Lab filter
- Gonen Lab (32) Apply Gonen Lab filter
- Grigorieff Lab (28) Apply Grigorieff Lab filter
- Harris Lab (10) Apply Harris Lab filter
- Heberlein Lab (81) Apply Heberlein Lab filter
- Hermundstad Lab (4) Apply Hermundstad Lab filter
- Hess Lab (3) Apply Hess Lab filter
- Jayaraman Lab (4) Apply Jayaraman Lab filter
- Johnson Lab (5) Apply Johnson Lab filter
- Kainmueller Lab (19) Apply Kainmueller Lab filter
- Karpova Lab (1) Apply Karpova Lab filter
- Keleman Lab (5) Apply Keleman Lab filter
- Keller Lab (15) Apply Keller Lab filter
- Koay Lab (16) Apply Koay Lab filter
- Lavis Lab (12) Apply Lavis Lab filter
- Lee (Albert) Lab (5) Apply Lee (Albert) Lab filter
- Leonardo Lab (4) Apply Leonardo Lab filter
- Li Lab (24) Apply Li Lab filter
- Lippincott-Schwartz Lab (72) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (5) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (5) Apply Liu (Zhe) Lab filter
- Looger Lab (1) Apply Looger Lab filter
- Magee Lab (18) Apply Magee Lab filter
- Menon Lab (6) Apply Menon Lab filter
- Murphy Lab (7) Apply Murphy Lab filter
- O'Shea Lab (1) Apply O'Shea Lab filter
- Otopalik Lab (12) Apply Otopalik Lab filter
- Pachitariu Lab (12) Apply Pachitariu Lab filter
- Pastalkova Lab (13) Apply Pastalkova Lab filter
- Pavlopoulos Lab (12) Apply Pavlopoulos Lab filter
- Pedram Lab (11) Apply Pedram Lab filter
- Reiser Lab (6) Apply Reiser Lab filter
- Riddiford Lab (24) Apply Riddiford Lab filter
- Romani Lab (12) Apply Romani Lab filter
- Rubin Lab (38) Apply Rubin Lab filter
- Saalfeld Lab (17) Apply Saalfeld Lab filter
- Satou Lab (15) Apply Satou Lab filter
- Schreiter Lab (17) Apply Schreiter Lab filter
- Sgro Lab (20) Apply Sgro Lab filter
- Simpson Lab (5) Apply Simpson Lab filter
- Singer Lab (43) Apply Singer Lab filter
- Spruston Lab (36) Apply Spruston Lab filter
- Stern Lab (83) Apply Stern Lab filter
- Sternson Lab (7) Apply Sternson Lab filter
- Stringer Lab (3) Apply Stringer Lab filter
- Svoboda Lab (4) Apply Svoboda Lab filter
- Tebo Lab (24) Apply Tebo Lab filter
- Tillberg Lab (3) Apply Tillberg Lab filter
- Tjian Lab (47) Apply Tjian Lab filter
- Truman Lab (30) Apply Truman Lab filter
- Turaga Lab (12) Apply Turaga Lab filter
- Turner Lab (11) Apply Turner Lab filter
- Wang (Shaohe) Lab (19) Apply Wang (Shaohe) Lab filter
- Wu Lab (1) Apply Wu Lab filter
- Zlatic Lab (2) Apply Zlatic Lab filter
- Zuker Lab (20) Apply Zuker Lab filter
Associated Project Team
Publication Date
- 2024 (1) Apply 2024 filter
- 2023 (1) Apply 2023 filter
- 2022 (26) Apply 2022 filter
- 2021 (19) Apply 2021 filter
- 2020 (19) Apply 2020 filter
- 2019 (25) Apply 2019 filter
- 2018 (26) Apply 2018 filter
- 2017 (31) Apply 2017 filter
- 2016 (18) Apply 2016 filter
- 2015 (57) Apply 2015 filter
- 2014 (46) Apply 2014 filter
- 2013 (58) Apply 2013 filter
- 2012 (78) Apply 2012 filter
- 2011 (92) Apply 2011 filter
- 2010 (100) Apply 2010 filter
- 2009 (102) Apply 2009 filter
- 2008 (100) Apply 2008 filter
- 2007 (85) Apply 2007 filter
- 2006 (89) Apply 2006 filter
- 2005 (67) Apply 2005 filter
- 2004 (57) Apply 2004 filter
- 2003 (58) Apply 2003 filter
- 2002 (39) Apply 2002 filter
- 2001 (28) Apply 2001 filter
- 2000 (29) Apply 2000 filter
- 1999 (14) Apply 1999 filter
- 1998 (18) Apply 1998 filter
- 1997 (16) Apply 1997 filter
- 1996 (10) Apply 1996 filter
- 1995 (18) Apply 1995 filter
- 1994 (12) Apply 1994 filter
- 1993 (10) Apply 1993 filter
- 1992 (6) Apply 1992 filter
- 1991 (11) Apply 1991 filter
- 1990 (11) Apply 1990 filter
- 1989 (6) Apply 1989 filter
- 1988 (1) Apply 1988 filter
- 1987 (7) Apply 1987 filter
- 1986 (4) Apply 1986 filter
- 1985 (5) Apply 1985 filter
- 1984 (2) Apply 1984 filter
- 1983 (2) Apply 1983 filter
- 1982 (3) Apply 1982 filter
- 1981 (3) Apply 1981 filter
- 1980 (1) Apply 1980 filter
- 1979 (1) Apply 1979 filter
- 1976 (2) Apply 1976 filter
- 1973 (1) Apply 1973 filter
- 1970 (1) Apply 1970 filter
- 1967 (1) Apply 1967 filter
Type of Publication
- Remove Non-Janelia filter Non-Janelia
1417 Publications
Showing 1251-1260 of 1417 resultsPURPOSE: To discover proteins that have the potential to contribute to the tight packing of fiber cells in the lens. METHODS: Crude fiber cell membranes were isolated from ovine lens cortex. Proteins were separated by two-dimensional gel electrophoresis, and selected protein spots identified by micro-sequencing. The identification of galectin-3 was confirmed by immunoblotting with a specific antibody. The association of galectin-3 with the fiber cell plasma membrane was investigated using immunofluorescence microscopy, solubilization trials with selected reagents, and immunoprecipitation to identify candidate ligands. RESULTS: A cluster of three protein spots with an apparent molecular weight of 31,000 and isoelectric points ranging between 7 and 8.5 were resolved and identified as galectin-3. This protein was associated peripherally with the fiber cell plasma membrane and interacted with MP20, an abundant intrinsic membrane protein that had been identified previously as a component of membrane junctions between fiber cells. CONCLUSIONS: The detection of galectin-3 in the lens is a novel result and adds to the growing list of lens proteins with adhesive properties. Its location at the fiber cell membrane and its association with the junction-forming MP20 is consistent with a potential role in the development or maintenance of the tightly packed lens tissue architecture.
In CA1 pyramidal neurons of the hippocampus, calcium-dependent spikes occur in vivo during specific behavioral states and may be enhanced during epileptiform activity. However, the mechanisms that control calcium spike initiation and repolarization are poorly understood. Using dendritic and somatic patch-pipette recordings, we show that calcium spikes are initiated in the apical dendrites of CA1 pyramidal neurons and drive bursts of sodium-dependent action potentials at the soma. Initiation of calcium spikes at the soma was suppressed in part by potassium channels activated by sodium-dependent action potentials. Low-threshold, putative D-type potassium channels [blocked by 100 microM 4-aminopyridine (4-AP) and 0.5-1 microM alpha-dendrotoxin (alpha-DTX)] played a prominent role in setting a high threshold for somatic calcium spikes, thus restricting initiation to the dendrites. DTX- and 4-AP-sensitive channels were activated during sodium-dependent action potentials and mediated a large component of their afterhyperpolarization. Once initiated, repetitive firing of calcium spikes was limited by activation of putative BK-type calcium-activated potassium channels (blocked by 250 microM tetraethylammonium chloride, 70 nM charybdotoxin, or 100 nM iberiotoxin). Thus, the concerted action of calcium- and voltage-activated potassium channels serves to focus spatially and temporally the membrane depolarization and calcium influx generated by calcium spikes during strong, synchronous network excitation.
Insect metamorphosis is a fascinating and highly successful biological adaptation, but there is much uncertainty as to how it evolved. Ancestral insect species did not undergo metamorphosis and there are still some existing species that lack metamorphosis or undergo only partial metamorphosis. Based on endocrine studies and morphological comparisons of the development of insect species with and without metamorphosis, a novel hypothesis for the evolution of metamorphosis is proposed. Changes in the endocrinology of development are central to this hypothesis. The three stages of the ancestral insect species-pronymph, nymph and adult-are proposed to be equivalent to the larva, pupa and adult stages of insects with complete metamorphosis. This proposal has general implications for insect developmental biology.
The mushroom bodies (MBs) are prominent structures in the Drosophila brain that are essential for olfactory learning and memory. Characterization of the development and projection patterns of individual MB neurons will be important for elucidating their functions. Using mosaic analysis with a repressible cell marker (Lee, T. and Luo, L. (1999) Neuron 22, 451-461), we have positively marked the axons and dendrites of multicellular and single-cell mushroom body clones at specific developmental stages. Systematic clonal analysis demonstrates that a single mushroom body neuroblast sequentially generates at least three types of morphologically distinct neurons. Neurons projecting into the (gamma) lobe of the adult MB are born first, prior to the mid-3rd instar larval stage. Neurons projecting into the alpha’ and beta’ lobes are born between the mid-3rd instar larval stage and puparium formation. Finally, neurons projecting into the alpha and beta lobes are born after puparium formation. Visualization of individual MB neurons has also revealed how different neurons acquire their characteristic axon projections. During the larval stage, axons of all MB neurons bifurcate into both the dorsal and medial lobes. Shortly after puparium formation, larval MB neurons are selectively pruned according to birthdays. Degeneration of axon branches makes early-born gamma neurons retain only their main processes in the peduncle, which then project into the adult gamma lobe without bifurcation. In contrast, the basic axon projections of the later-born (alpha’/beta’) larval neurons are preserved during metamorphosis. This study illustrates the cellular organization of mushroom bodies and the development of different MB neurons at the single cell level. It allows for future studies on the molecular mechanisms of mushroom body development.
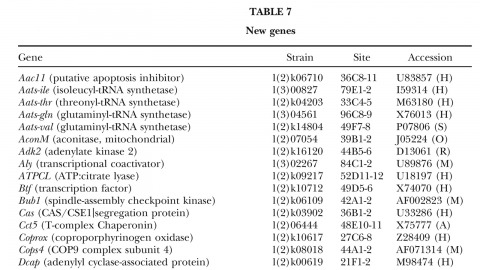
A fundamental goal of genetics and functional genomics is to identify and mutate every gene in model organisms such as Drosophila melanogaster. The Berkeley Drosophila Genome Project (BDGP) gene disruption project generates single P-element insertion strains that each mutate unique genomic open reading frames. Such strains strongly facilitate further genetic and molecular studies of the disrupted loci, but it has remained unclear if P elements can be used to mutate all Drosophila genes. We now report that the primary collection has grown to contain 1045 strains that disrupt more than 25% of the estimated 3600 Drosophila genes that are essential for adult viability. Of these P insertions, 67% have been verified by genetic tests to cause the associated recessive mutant phenotypes, and the validity of most of the remaining lines is predicted on statistical grounds. Sequences flanking >920 insertions have been determined to exactly position them in the genome and to identify 376 potentially affected transcripts from collections of EST sequences. Strains in the BDGP collection are available from the Bloomington Stock Center and have already assisted the research community in characterizing >250 Drosophila genes. The likely identity of 131 additional genes in the collection is reported here. Our results show that Drosophila genes have a wide range of sensitivity to inactivation by P elements, and provide a rationale for greatly expanding the BDGP primary collection based entirely on insertion site sequencing. We predict that this approach can bring >85% of all Drosophila open reading frames under experimental control.
A room-temperature molten salt has been prepared from AuCl3 and 1-ethyl-3-methylimidazolium chloride (EMIC). At a ratio of 1 mol of AuCl3 to 2 mol of EMIC, the salt is a bright yellow-orange and shows Raman spectral features at 170, 328, and 352 cm-1, indicating the presence of AuCl4-. Ab initio calculations indicate that a dinuclear Au2Cl7- species containing a bridging chlorine should be stable, but no such species has been observed.
Young birds learn to sing by using auditory feedback to compare their own vocalizations to a memorized or innate song pattern; if they are deafened as juveniles, they will not develop normal songs. The completion of song development is called crystallization. After this stage, song shows little variation in its temporal or spectral properties. However, the mechanisms underlying this stability are largely unknown. Here we present evidence that auditory feedback is actively used in adulthood to maintain the stability of song structure. We found that perturbing auditory feedback during singing in adult zebra finches caused their song to deteriorate slowly. This ’decrystallization’ consisted of a marked loss of the spectral and temporal stereotypy seen in crystallized song, including stuttering, creation, deletion and distortion of song syllables. After normal feedback was restored, these deviations gradually disappeared and the original song was recovered. Thus, adult birds that do not learn new songs nevertheless retain a significant amount of plasticity in the brain.
We describe a genetic mosaic system in Drosophila, in which a dominant repressor of a cell marker is placed in trans to a mutant gene of interest. Mitotic recombination events between homologous chromosomes generate homozygous mutant cells, which are exclusively labeled due to loss of the repressor. Using this system, we are able to visualize axonal projections and dendritic elaboration in large neuroblast clones and single neuron clones with a membrane-targeted GFP marker. This new method allows for the study of gene functions in neuroblast proliferation, axon guidance, and dendritic elaboration in the complex central nervous system. As an example, we show that the short stop gene is required in mushroom body neurons for the extension and guidance of their axons.