Filter
Associated Lab
- Aguilera Castrejon Lab (1) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (2) Apply Ahrens Lab filter
- Aso Lab (3) Apply Aso Lab filter
- Baker Lab (1) Apply Baker Lab filter
- Betzig Lab (8) Apply Betzig Lab filter
- Beyene Lab (2) Apply Beyene Lab filter
- Bock Lab (1) Apply Bock Lab filter
- Branson Lab (7) Apply Branson Lab filter
- Card Lab (4) Apply Card Lab filter
- Cardona Lab (8) Apply Cardona Lab filter
- Cui Lab (1) Apply Cui Lab filter
- Darshan Lab (1) Apply Darshan Lab filter
- Dickson Lab (1) Apply Dickson Lab filter
- Druckmann Lab (3) Apply Druckmann Lab filter
- Dudman Lab (4) Apply Dudman Lab filter
- Eddy/Rivas Lab (1) Apply Eddy/Rivas Lab filter
- Feliciano Lab (1) Apply Feliciano Lab filter
- Fetter Lab (4) Apply Fetter Lab filter
- Funke Lab (1) Apply Funke Lab filter
- Gonen Lab (11) Apply Gonen Lab filter
- Grigorieff Lab (6) Apply Grigorieff Lab filter
- Harris Lab (5) Apply Harris Lab filter
- Heberlein Lab (1) Apply Heberlein Lab filter
- Hermundstad Lab (1) Apply Hermundstad Lab filter
- Hess Lab (4) Apply Hess Lab filter
- Jayaraman Lab (4) Apply Jayaraman Lab filter
- Ji Lab (5) Apply Ji Lab filter
- Johnson Lab (1) Apply Johnson Lab filter
- Kainmueller Lab (1) Apply Kainmueller Lab filter
- Keleman Lab (1) Apply Keleman Lab filter
- Keller Lab (2) Apply Keller Lab filter
- Koay Lab (3) Apply Koay Lab filter
- Lavis Lab (16) Apply Lavis Lab filter
- Lee (Albert) Lab (6) Apply Lee (Albert) Lab filter
- Leonardo Lab (2) Apply Leonardo Lab filter
- Li Lab (4) Apply Li Lab filter
- Lippincott-Schwartz Lab (11) Apply Lippincott-Schwartz Lab filter
- Liu (Zhe) Lab (5) Apply Liu (Zhe) Lab filter
- Looger Lab (6) Apply Looger Lab filter
- Magee Lab (2) Apply Magee Lab filter
- Menon Lab (1) Apply Menon Lab filter
- Otopalik Lab (3) Apply Otopalik Lab filter
- Pachitariu Lab (5) Apply Pachitariu Lab filter
- Pedram Lab (1) Apply Pedram Lab filter
- Reiser Lab (6) Apply Reiser Lab filter
- Riddiford Lab (1) Apply Riddiford Lab filter
- Romani Lab (6) Apply Romani Lab filter
- Rubin Lab (15) Apply Rubin Lab filter
- Saalfeld Lab (4) Apply Saalfeld Lab filter
- Scheffer Lab (4) Apply Scheffer Lab filter
- Schreiter Lab (4) Apply Schreiter Lab filter
- Shroff Lab (1) Apply Shroff Lab filter
- Simpson Lab (2) Apply Simpson Lab filter
- Singer Lab (6) Apply Singer Lab filter
- Spruston Lab (1) Apply Spruston Lab filter
- Stern Lab (8) Apply Stern Lab filter
- Sternson Lab (2) Apply Sternson Lab filter
- Svoboda Lab (9) Apply Svoboda Lab filter
- Tebo Lab (3) Apply Tebo Lab filter
- Truman Lab (6) Apply Truman Lab filter
- Turaga Lab (3) Apply Turaga Lab filter
- Turner Lab (2) Apply Turner Lab filter
- Wang (Shaohe) Lab (4) Apply Wang (Shaohe) Lab filter
- Wu Lab (1) Apply Wu Lab filter
- Zlatic Lab (7) Apply Zlatic Lab filter
Associated Project Team
- Fly Descending Interneuron (1) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (4) Apply Fly Functional Connectome filter
- Fly Olympiad (1) Apply Fly Olympiad filter
- FlyEM (4) Apply FlyEM filter
- FlyLight (2) Apply FlyLight filter
- GENIE (3) Apply GENIE filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (3) Apply Tool Translation Team (T3) filter
- Transcription Imaging (6) Apply Transcription Imaging filter
Publication Date
- December 2017 (16) Apply December 2017 filter
- November 2017 (13) Apply November 2017 filter
- October 2017 (10) Apply October 2017 filter
- September 2017 (14) Apply September 2017 filter
- August 2017 (18) Apply August 2017 filter
- July 2017 (25) Apply July 2017 filter
- June 2017 (19) Apply June 2017 filter
- May 2017 (27) Apply May 2017 filter
- April 2017 (22) Apply April 2017 filter
- March 2017 (19) Apply March 2017 filter
- February 2017 (13) Apply February 2017 filter
- January 2017 (21) Apply January 2017 filter
- Remove 2017 filter 2017
Type of Publication
217 Publications
Showing 31-40 of 217 results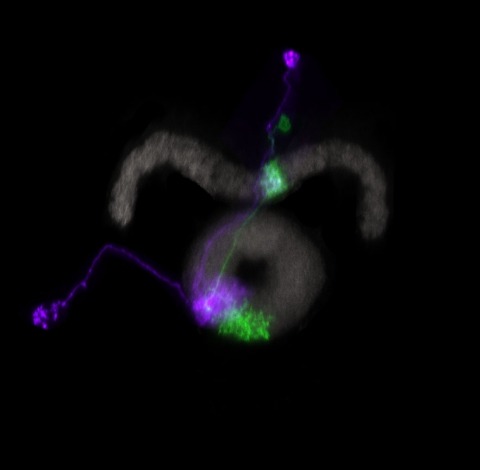
Many animals maintain an internal representation of their heading as they move through their surroundings. Such a compass representation was recently discovered in a neural population in the Drosophila melanogaster central complex, a brain region implicated in spatial navigation. Here, we use two-photon calcium imaging and electrophysiology in head-fixed walking flies to identify a different neural population that conjunctively encodes heading and angular velocity, and is excited selectively by turns in either the clockwise or counterclockwise direction. We show how these mirror-symmetric turn responses combine with the neurons' connectivity to the compass neurons to create an elegant mechanism for updating the fly's heading representation when the animal turns in darkness. This mechanism, which employs recurrent loops with an angular shift, bears a resemblance to those proposed in theoretical models for rodent head direction cells. Our results provide a striking example of structure matching function for a broadly relevant computation.
Although purification of biotinylated molecules is highly efficient, identifying specific sites of biotinylation remains challenging. We show that anti-biotin antibodies enable unprecedented enrichment of biotinylated peptides from complex peptide mixtures. Live-cell proximity labeling using APEX peroxidase followed by anti-biotin enrichment and mass spectrometry yielded over 1,600 biotinylation sites on hundreds of proteins, an increase of more than 30-fold in the number of biotinylation sites identified compared to streptavidin-based enrichment of proteins.
The organization of the eukaryotic cell into discrete membrane-bound organelles allows for the separation of incompatible biochemical processes, but the activities of these organelles must be coordinated. For example, lipid metabolism is distributed between the endoplasmic reticulum for lipid synthesis, lipid droplets for storage and transport, mitochondria and peroxisomes for β-oxidation, and lysosomes for lipid hydrolysis and recycling. It is increasingly recognized that organelle contacts have a vital role in diverse cellular functions. However, the spatial and temporal organization of organelles within the cell remains poorly characterized, as fluorescence imaging approaches are limited in the number of different labels that can be distinguished in a single image. Here we present a systems-level analysis of the organelle interactome using a multispectral image acquisition method that overcomes the challenge of spectral overlap in the fluorescent protein palette. We used confocal and lattice light sheet instrumentation and an imaging informatics pipeline of five steps to achieve mapping of organelle numbers, volumes, speeds, positions and dynamic inter-organelle contacts in live cells from a monkey fibroblast cell line. We describe the frequency and locality of two-, three-, four- and five-way interactions among six different membrane-bound organelles (endoplasmic reticulum, Golgi, lysosome, peroxisome, mitochondria and lipid droplet) and show how these relationships change over time. We demonstrate that each organelle has a characteristic distribution and dispersion pattern in three-dimensional space and that there is a reproducible pattern of contacts among the six organelles, that is affected by microtubule and cell nutrient status. These live-cell confocal and lattice light sheet spectral imaging approaches are applicable to any cell system expressing multiple fluorescent probes, whether in normal conditions or when cells are exposed to disturbances such as drugs, pathogens or stress. This methodology thus offers a powerful descriptive tool and can be used to develop hypotheses about cellular organization and dynamics.
hIAPP fibrils are associated with Type-II Diabetes, but the link of hIAPP structure to islet cell death remains elusive. Here we observe that hIAPP fibrils are cytotoxic to cultured pancreatic β-cells, leading us to determine the structure and cytotoxicity of protein segments composing the amyloid spine of hIAPP. Using the cryoEM method MicroED, we discover that one segment, 19-29 S20G, forms pairs of β-sheets mated by a dry interface that share structural features with and are similarly cytotoxic to full-length hIAPP fibrils. In contrast, a second segment, 15-25 WT, forms non-toxic labile β-sheets. These segments possess different structures and cytotoxic effects, however, both can seed full-length hIAPP, and cause hIAPP to take on the cytotoxic and structural features of that segment. These results suggest that protein segment structures represent polymorphs of their parent protein and that segment 19-29 S20G may serve as a model for the toxic spine of hIAPP.
Traditionally, crystallographic analysis of macromolecules has depended on large, well-ordered crystals, which often require significant effort to obtain. Even sizable crystals sometimes suffer from pathologies that render them inappropriate for high-resolution structure determination. Here we show that fragmentation of large, imperfect crystals into microcrystals or nanocrystals can provide a simple path for high-resolution structure determination by the cryoEM method MicroED and potentially by serial femtosecond crystallography.
Although the endoplasmic reticulum (ER) extends throughout axons and axonal ER dysfunction is implicated in numerous neurological diseases, its role at nerve terminals is poorly understood. We developed novel genetically encoded ER-targeted low-affinity Ca(2+) indicators optimized for examining axonal ER Ca(2+). Our experiments revealed that presynaptic function is tightly controlled by ER Ca(2+) content. We found that neuronal activity drives net Ca(2+) uptake into presynaptic ER although this activity does not contribute significantly to shaping cytosolic Ca(2+) except during prolonged repetitive firing. In contrast, we found that axonal ER acts as an actuator of plasma membrane (PM) function: [Ca(2+)]ER controls STIM1 activation in presynaptic terminals, which results in the local modulation of presynaptic function, impacting activity-driven Ca(2+) entry and release probability. These experiments reveal a critical role of presynaptic ER in the control of neurotransmitter release and will help frame future investigations into the molecular basis of ER-driven neuronal disease states.
The behavioral state of an animal can dynamically modulate visual processing. In flies, the behavioral state is known to alter the temporal tuning of neurons that carry visual motion information into the central brain. However, where this modulation occurs and how it tunes the properties of this neural circuit are not well understood. Here, we show that the behavioral state alters the baseline activity levels and the temporal tuning of the first directionally selective neuron in the ON motion pathway (T4) as well as its primary input neurons (Mi1, Tm3, Mi4, Mi9). These effects are especially prominent in the inhibitory neuron Mi4, and we show that central octopaminergic neurons provide input to Mi4 and increase its excitability. We further show that octopamine neurons are required for sustained behavioral responses to fast-moving, but not slow-moving, visual stimuli in walking flies. These results indicate that behavioral-state modulation acts directly on the inputs to the directionally selective neurons and supports efficient neural coding of motion stimuli.
Learning is primarily mediated by activity-dependent modifications of synaptic strength within neuronal circuits. We discovered that place fields in hippocampal area CA1 are produced by a synaptic potentiation notably different from Hebbian plasticity. Place fields could be produced in vivo in a single trial by potentiation of input that arrived seconds before and after complex spiking. The potentiated synaptic input was not initially coincident with action potentials or depolarization. This rule, named behavioral time scale synaptic plasticity, abruptly modifies inputs that were neither causal nor close in time to postsynaptic activation. In slices, five pairings of subthreshold presynaptic activity and calcium (Ca(2+)) plateau potentials produced a large potentiation with an asymmetric seconds-long time course. This plasticity efficiently stores entire behavioral sequences within synaptic weights to produce predictive place cell activity.
This paper provides an overview of the discussion and presentations from the Workshop on the Management of Large CryoEM Facilities held at the New York Structural Biology Center, New York, NY on February 6–7, 2017. A major objective of the workshop was to discuss best practices for managing cryoEM facilities. The discussions were largely focused on supporting single-particle methods for cryoEM and topics included: user access, assessing projects, workflow, sample handling, microscopy, data management and processing, and user training.
While building information modeling (BIM) is widely embraced by the architectural, engineering and construction (AEC) industry, BIM adoption in facilities management (FM) is still relatively new and limited. BIM deliverables from design and construction generally do not fulfill FM needs unless they are clearly specified and carefully managed. The Facilities Group responsible for the Janelia Research Campus of the Howard Hughes Medical Institute (HHMI) expects any BIM platform to provide value in operations and maintenance. Janelia’s BIM vision goes beyond transferring BIM data to computerized maintenance management software (CMMS) and integrated workplace management system (IWMS) platforms. Instead, Janelia creates and maintains FM-capable BIM, utilizes the models to solve operational challenges and improves safety and efficiency in various ways, including engineering analysis for heating, ventilation and air conditioning (HVAC), electrical and plumbing; building automation systems (BAS) analysis; operational impact analysis; and BIM-aided operation safety.