Filter
Associated Lab
- Ahrens Lab (2) Apply Ahrens Lab filter
- Aso Lab (3) Apply Aso Lab filter
- Baker Lab (1) Apply Baker Lab filter
- Betzig Lab (8) Apply Betzig Lab filter
- Bock Lab (1) Apply Bock Lab filter
- Branson Lab (7) Apply Branson Lab filter
- Card Lab (4) Apply Card Lab filter
- Cardona Lab (8) Apply Cardona Lab filter
- Cui Lab (1) Apply Cui Lab filter
- Dickson Lab (1) Apply Dickson Lab filter
- Druckmann Lab (3) Apply Druckmann Lab filter
- Dudman Lab (4) Apply Dudman Lab filter
- Eddy/Rivas Lab (1) Apply Eddy/Rivas Lab filter
- Feliciano Lab (1) Apply Feliciano Lab filter
- Fetter Lab (4) Apply Fetter Lab filter
- Funke Lab (1) Apply Funke Lab filter
- Gonen Lab (11) Apply Gonen Lab filter
- Grigorieff Lab (6) Apply Grigorieff Lab filter
- Harris Lab (5) Apply Harris Lab filter
- Heberlein Lab (1) Apply Heberlein Lab filter
- Hermundstad Lab (1) Apply Hermundstad Lab filter
- Hess Lab (4) Apply Hess Lab filter
- Jayaraman Lab (4) Apply Jayaraman Lab filter
- Ji Lab (5) Apply Ji Lab filter
- Keleman Lab (1) Apply Keleman Lab filter
- Keller Lab (2) Apply Keller Lab filter
- Lavis Lab (16) Apply Lavis Lab filter
- Lee (Albert) Lab (6) Apply Lee (Albert) Lab filter
- Leonardo Lab (2) Apply Leonardo Lab filter
- Lippincott-Schwartz Lab (9) Apply Lippincott-Schwartz Lab filter
- Liu (Zhe) Lab (5) Apply Liu (Zhe) Lab filter
- Looger Lab (6) Apply Looger Lab filter
- Magee Lab (2) Apply Magee Lab filter
- Menon Lab (1) Apply Menon Lab filter
- Pachitariu Lab (1) Apply Pachitariu Lab filter
- Reiser Lab (6) Apply Reiser Lab filter
- Riddiford Lab (1) Apply Riddiford Lab filter
- Romani Lab (6) Apply Romani Lab filter
- Rubin Lab (15) Apply Rubin Lab filter
- Saalfeld Lab (4) Apply Saalfeld Lab filter
- Scheffer Lab (4) Apply Scheffer Lab filter
- Schreiter Lab (4) Apply Schreiter Lab filter
- Shroff Lab (1) Apply Shroff Lab filter
- Simpson Lab (2) Apply Simpson Lab filter
- Singer Lab (6) Apply Singer Lab filter
- Spruston Lab (1) Apply Spruston Lab filter
- Stern Lab (8) Apply Stern Lab filter
- Sternson Lab (2) Apply Sternson Lab filter
- Svoboda Lab (9) Apply Svoboda Lab filter
- Truman Lab (6) Apply Truman Lab filter
- Turaga Lab (3) Apply Turaga Lab filter
- Turner Lab (2) Apply Turner Lab filter
- Wu Lab (1) Apply Wu Lab filter
- Zlatic Lab (7) Apply Zlatic Lab filter
Associated Project Team
- Fly Descending Interneuron (1) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (4) Apply Fly Functional Connectome filter
- Fly Olympiad (1) Apply Fly Olympiad filter
- FlyEM (4) Apply FlyEM filter
- FlyLight (2) Apply FlyLight filter
- GENIE (3) Apply GENIE filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (3) Apply Tool Translation Team (T3) filter
- Transcription Imaging (6) Apply Transcription Imaging filter
Associated Support Team
- Anatomy and Histology (2) Apply Anatomy and Histology filter
- Cryo-Electron Microscopy (4) Apply Cryo-Electron Microscopy filter
- Electron Microscopy (1) Apply Electron Microscopy filter
- Fly Facility (1) Apply Fly Facility filter
- Integrative Imaging (1) Apply Integrative Imaging filter
- Project Technical Resources (1) Apply Project Technical Resources filter
- Quantitative Genomics (2) Apply Quantitative Genomics filter
- Scientific Computing Software (9) Apply Scientific Computing Software filter
- Viral Tools (1) Apply Viral Tools filter
- Vivarium (1) Apply Vivarium filter
Publication Date
- December 2017 (15) Apply December 2017 filter
- November 2017 (11) Apply November 2017 filter
- October 2017 (7) Apply October 2017 filter
- September 2017 (14) Apply September 2017 filter
- August 2017 (15) Apply August 2017 filter
- July 2017 (20) Apply July 2017 filter
- June 2017 (18) Apply June 2017 filter
- May 2017 (25) Apply May 2017 filter
- April 2017 (21) Apply April 2017 filter
- March 2017 (15) Apply March 2017 filter
- February 2017 (7) Apply February 2017 filter
- January 2017 (18) Apply January 2017 filter
- Remove 2017 filter 2017
186 Janelia Publications
Showing 21-30 of 186 resultsThe past quarter century has witnessed rapid developments of fluorescence microscopy techniques that enable structural and functional imaging of biological specimens at unprecedented depth and resolution. The performance of these methods in multicellular organisms, however, is degraded by sample-induced optical aberrations. Here I review recent work on incorporating adaptive optics, a technology originally applied in astronomical telescopes to combat atmospheric aberrations, to improve image quality of fluorescence microscopy for biological imaging.
Adjusting the objective correction collar is a widely used approach to correct spherical aberrations (SA) in optical microscopy. In this work, we characterized and compared its performance with adaptive optics in the context of in vivo brain imaging with two-photon fluorescence microscopy. We found that the presence of sample tilt had a deleterious effect on the performance of SA-only correction. At large tilt angles, adjusting the correction collar even worsened image quality. In contrast, adaptive optical correction always recovered optimal imaging performance regardless of sample tilt. The extent of improvement with adaptive optics was dependent on object size, with smaller objects having larger relative gains in signal intensity and image sharpness. These observations translate into a superior performance of adaptive optics for structural and functional brain imaging applications in vivo, as we confirmed experimentally.
The calcium-modulated photoactivatable ratiometric integrator CaMPARI (Fosque et al., 2015) facilitates the study of neural circuits by permanently marking cells active during user-specified temporal windows. Permanent marking enables measurement of signals from large swathes of tissue and easy correlation of activity with other structural or functional labels. One potential application of CaMPARI is labeling neurons postsynaptic to specific populations targeted for optogenetic stimulation, giving rise to all-optical functional connectivity mapping. Here, we characterized the response of CaMPARI to several common types of neuronal calcium signals in mouse acute cortical brain slices. Our experiments show that CaMPARI is effectively converted by both action potentials and sub-threshold synaptic inputs, and that conversion level is correlated to synaptic strength. Importantly, we found that conversion rate can be tuned: it is linearly related to light intensity. At low photoconversion light levels CaMPARI offers a wide dynamic range due to slower conversion rate; at high light levels conversion is more rapid and more sensitive to activity. Finally, we employed CaMPARI and optogenetics for functional circuit mapping in ex vivo acute brain slices, which preserve in vivo-like connectivity of axon terminals. With a single light source, we stimulated channelrhodopsin-2-expressing long-range posteromedial (POm) thalamic axon terminals in cortex and induced CaMPARI conversion in recipient cortical neurons. We found that POm stimulation triggers robust photoconversion of layer 5 cortical neurons and weaker conversion of layer 2/3 neurons. Thus, CaMPARI enables network-wide, tunable, all-optical functional circuit mapping that captures supra- and sub-threshold depolarization. This article is protected by copyright. All rights reserved.
Dietary restriction increases the longevity of many organisms but the cell signaling and organellar mechanisms underlying this capability are unclear. We demonstrate that to permit long-term survival in response to sudden glucose depletion, yeast cells activate lipid-droplet (LD) consumption through micro-lipophagy (µ-lipophagy), in which fat is metabolized as an alternative energy source. AMP-activated protein kinase (AMPK) activation triggered this pathway, which required Atg14p. More gradual glucose starvation, amino acid deprivation or rapamycin did not trigger µ-lipophagy and failed to provide the needed substitute energy source for long-term survival. During acute glucose restriction, activated AMPK was stabilized from degradation and interacted with Atg14p. This prompted Atg14p redistribution from ER exit sites onto liquid-ordered vacuole membrane domains, initiating µ-lipophagy. Our findings that activated AMPK and Atg14p are required to orchestrate µ-lipophagy for energy production in starved cells is relevant for studies on aging and evolutionary survival strategies of different organisms.
Mycobacteria possess a multi-layered cell wall that requires extensive remodelling during cell division. We investigated the role of an amidase_3 domain-containing N-acetylmuramyl-L-alanine amidase, a peptidoglycan remodelling enzyme implicated in cell division. We demonstrated that deletion of MSMEG_6281 (Ami1) in Mycobacterium smegmatis resulted in the formation of cellular chains, illustrative of cells that were unable to complete division. Suprisingly, viability in the Δami1 mutant was maintained through atypical lateral branching, the products of which proceeded to form viable daughter cells. We showed that these lateral buds resulted from mislocalization of DivIVA, a major determinant in facilitating polar elongation in mycobacterial cells. Failure of Δami1 mutant cells to separate also led to dysregulation of FtsZ ring bundling. Loss of Ami1 resulted in defects in septal peptidoglycan turnover with release of excess cell wall material from the septum or newly born cell poles. We noted signficant accumulation of 3-3 crosslinked muropeptides in the Δami1 mutant. We further demonstrated that deletion of ami1 leads to increased cell wall permeability and enhanced susceptiblity to cell wall targeting antibiotics. Collectively, these data provide novel insight on cell division in actinobacteria and highlights a new class of potential drug targets for mycobacterial diseases.
Behaviors involving the interaction of multiple individuals are complex and frequently crucial for an animal's survival. These interactions, ranging across sensory modalities, length scales, and time scales, are often subtle and difficult to characterize. Contextual effects on the frequency of behaviors become even more difficult to quantify when physical interaction between animals interferes with conventional data analysis, e.g. due to visual occlusion. We introduce a method for quantifying behavior in fruit fly interaction that combines high-throughput video acquisition and tracking of individuals with recent unsupervised methods for capturing an animal's entire behavioral repertoire. We find behavioral differences between solitary flies and those paired with an individual of the opposite sex, identifying specific behaviors that are affected by social and spatial context. Our pipeline allows for a comprehensive description of the interaction between two individuals using unsupervised machine learning methods, and will be used to answer questions about the depth of complexity and variance in fruit fly courtship.
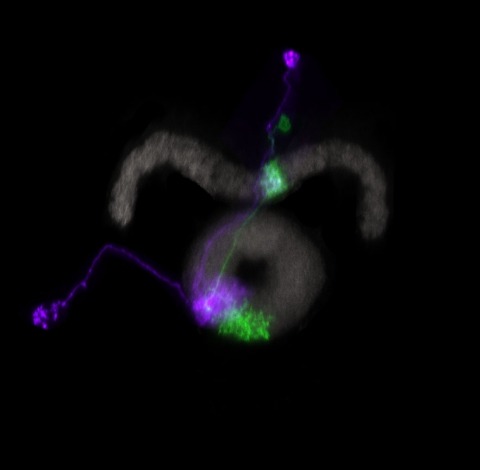
Many animals maintain an internal representation of their heading as they move through their surroundings. Such a compass representation was recently discovered in a neural population in the Drosophila melanogaster central complex, a brain region implicated in spatial navigation. Here, we use two-photon calcium imaging and electrophysiology in head-fixed walking flies to identify a different neural population that conjunctively encodes heading and angular velocity, and is excited selectively by turns in either the clockwise or counterclockwise direction. We show how these mirror-symmetric turn responses combine with the neurons' connectivity to the compass neurons to create an elegant mechanism for updating the fly's heading representation when the animal turns in darkness. This mechanism, which employs recurrent loops with an angular shift, bears a resemblance to those proposed in theoretical models for rodent head direction cells. Our results provide a striking example of structure matching function for a broadly relevant computation.
The organization of the eukaryotic cell into discrete membrane-bound organelles allows for the separation of incompatible biochemical processes, but the activities of these organelles must be coordinated. For example, lipid metabolism is distributed between the endoplasmic reticulum for lipid synthesis, lipid droplets for storage and transport, mitochondria and peroxisomes for β-oxidation, and lysosomes for lipid hydrolysis and recycling. It is increasingly recognized that organelle contacts have a vital role in diverse cellular functions. However, the spatial and temporal organization of organelles within the cell remains poorly characterized, as fluorescence imaging approaches are limited in the number of different labels that can be distinguished in a single image. Here we present a systems-level analysis of the organelle interactome using a multispectral image acquisition method that overcomes the challenge of spectral overlap in the fluorescent protein palette. We used confocal and lattice light sheet instrumentation and an imaging informatics pipeline of five steps to achieve mapping of organelle numbers, volumes, speeds, positions and dynamic inter-organelle contacts in live cells from a monkey fibroblast cell line. We describe the frequency and locality of two-, three-, four- and five-way interactions among six different membrane-bound organelles (endoplasmic reticulum, Golgi, lysosome, peroxisome, mitochondria and lipid droplet) and show how these relationships change over time. We demonstrate that each organelle has a characteristic distribution and dispersion pattern in three-dimensional space and that there is a reproducible pattern of contacts among the six organelles, that is affected by microtubule and cell nutrient status. These live-cell confocal and lattice light sheet spectral imaging approaches are applicable to any cell system expressing multiple fluorescent probes, whether in normal conditions or when cells are exposed to disturbances such as drugs, pathogens or stress. This methodology thus offers a powerful descriptive tool and can be used to develop hypotheses about cellular organization and dynamics.
hIAPP fibrils are associated with Type-II Diabetes, but the link of hIAPP structure to islet cell death remains elusive. Here we observe that hIAPP fibrils are cytotoxic to cultured pancreatic β-cells, leading us to determine the structure and cytotoxicity of protein segments composing the amyloid spine of hIAPP. Using the cryoEM method MicroED, we discover that one segment, 19-29 S20G, forms pairs of β-sheets mated by a dry interface that share structural features with and are similarly cytotoxic to full-length hIAPP fibrils. In contrast, a second segment, 15-25 WT, forms non-toxic labile β-sheets. These segments possess different structures and cytotoxic effects, however, both can seed full-length hIAPP, and cause hIAPP to take on the cytotoxic and structural features of that segment. These results suggest that protein segment structures represent polymorphs of their parent protein and that segment 19-29 S20G may serve as a model for the toxic spine of hIAPP.