Filter
Associated Lab
- Aso Lab (2) Apply Aso Lab filter
- Baker Lab (2) Apply Baker Lab filter
- Betzig Lab (5) Apply Betzig Lab filter
- Bock Lab (1) Apply Bock Lab filter
- Branson Lab (3) Apply Branson Lab filter
- Card Lab (1) Apply Card Lab filter
- Cardona Lab (1) Apply Cardona Lab filter
- Chklovskii Lab (1) Apply Chklovskii Lab filter
- Cui Lab (6) Apply Cui Lab filter
- Druckmann Lab (2) Apply Druckmann Lab filter
- Eddy/Rivas Lab (1) Apply Eddy/Rivas Lab filter
- Fetter Lab (1) Apply Fetter Lab filter
- Gonen Lab (4) Apply Gonen Lab filter
- Harris Lab (3) Apply Harris Lab filter
- Heberlein Lab (1) Apply Heberlein Lab filter
- Hess Lab (3) Apply Hess Lab filter
- Jayaraman Lab (2) Apply Jayaraman Lab filter
- Ji Lab (2) Apply Ji Lab filter
- Karpova Lab (1) Apply Karpova Lab filter
- Keller Lab (3) Apply Keller Lab filter
- Lavis Lab (4) Apply Lavis Lab filter
- Lee (Albert) Lab (2) Apply Lee (Albert) Lab filter
- Leonardo Lab (2) Apply Leonardo Lab filter
- Looger Lab (13) Apply Looger Lab filter
- Magee Lab (6) Apply Magee Lab filter
- Pastalkova Lab (1) Apply Pastalkova Lab filter
- Pavlopoulos Lab (1) Apply Pavlopoulos Lab filter
- Reiser Lab (1) Apply Reiser Lab filter
- Riddiford Lab (1) Apply Riddiford Lab filter
- Rubin Lab (7) Apply Rubin Lab filter
- Saalfeld Lab (1) Apply Saalfeld Lab filter
- Scheffer Lab (3) Apply Scheffer Lab filter
- Schreiter Lab (2) Apply Schreiter Lab filter
- Simpson Lab (1) Apply Simpson Lab filter
- Spruston Lab (2) Apply Spruston Lab filter
- Sternson Lab (4) Apply Sternson Lab filter
- Svoboda Lab (9) Apply Svoboda Lab filter
- Tervo Lab (1) Apply Tervo Lab filter
- Tjian Lab (1) Apply Tjian Lab filter
- Truman Lab (3) Apply Truman Lab filter
Associated Project Team
Associated Support Team
Publication Date
- December 2012 (6) Apply December 2012 filter
- November 2012 (11) Apply November 2012 filter
- October 2012 (14) Apply October 2012 filter
- September 2012 (3) Apply September 2012 filter
- August 2012 (8) Apply August 2012 filter
- July 2012 (5) Apply July 2012 filter
- June 2012 (10) Apply June 2012 filter
- May 2012 (7) Apply May 2012 filter
- April 2012 (9) Apply April 2012 filter
- March 2012 (6) Apply March 2012 filter
- February 2012 (11) Apply February 2012 filter
- January 2012 (22) Apply January 2012 filter
- Remove 2012 filter 2012
112 Janelia Publications
Showing 61-70 of 112 resultsThe GFP reconstitution across synaptic partners (GRASP) technique, based on functional complementation between two nonfluorescent GFP fragments, can be used to detect the location of synapses quickly, accurately and with high spatial resolution. The method has been previously applied in the nematode and the fruit fly but requires substantial modification for use in the mammalian brain. We developed mammalian GRASP (mGRASP) by optimizing transmembrane split-GFP carriers for mammalian synapses. Using in silico protein design, we engineered chimeric synaptic mGRASP fragments that were efficiently delivered to synaptic locations and reconstituted GFP fluorescence in vivo. Furthermore, by integrating molecular and cellular approaches with a computational strategy for the three-dimensional reconstruction of neurons, we applied mGRASP to both long-range circuits and local microcircuits in the mouse hippocampus and thalamocortical regions, analyzing synaptic distribution in single neurons and in dendritic compartments.
The ability to automatically segment an image into distinct regions is a critical aspect in many visual processing applications. Because inaccuracies often exist in automatic segmentation, manual segmentation is necessary in some application domains to correct mistakes, such as required in the reconstruction of neuronal processes from microscopic images. The goal of the automated segmentation tool is traditionally to produce the highest-quality segmentation, where quality is measured by the similarity to actual ground truth, so as to minimize the volume of manual correction necessary. Manual correction is generally orders-of-magnitude more time consuming than automated segmentation, often making handling large images intractable. Therefore, we propose a more relevant goal: minimizing the turn-around time of automated/manual segmentation while attaining a level of similarity with ground truth. It is not always necessary to inspect every aspect of an image to generate a useful segmentation. As such, we propose a strategy to guide manual segmentation to the most uncertain parts of segmentation. Our contributions include 1) a probabilistic measure that evaluates segmentation without ground truth and 2) a methodology that leverages these probabilistic measures to significantly reduce manual correction while maintaining segmentation quality.
Cell recognition requires interactions through molecules located on cell surface. The insect homolog of Down syndrome cell adhesion molecule (Dscam) manifests huge molecular diversity in its extracellular domain. High-affinity Dscam-Dscam interactions only occur between isoforms that carry identical extracellular domains. Homophilic Dscam signaling can, thus, vary in strength depending on the compositions of Dscams present on the opposing cell surfaces. Dscam abundantly exists in the developing nervous system and governs arborization and proper elaboration of neurites. Notably, individual neurons may stochastically and dynamically express a small subset of Dscam isoforms such that any given neurite can be endowed with a unique repertoire of Dscams. This allows individual neurites to recognize their sister branches. Self-recognition leads to self-repulsion, ensuring divergent migration of sister processes. By contrast, weak homophilic Dscam interactions may promote fasciculation of neurites that express analogous, but not identical, Dscams. Differential Dscam binding may provide graded cell recognition that in turn governs complex neuronal morphogenesis.
The mechanisms linking sensation and action during learning are poorly understood. Layer 2/3 neurons in the motor cortex might participate in sensorimotor integration and learning; they receive input from sensory cortex and excite deep layer neurons, which control movement. Here we imaged activity in the same set of layer 2/3 neurons in the motor cortex over weeks, while mice learned to detect objects with their whiskers and report detection with licking. Spatially intermingled neurons represented sensory (touch) and motor behaviours (whisker movements and licking). With learning, the population-level representation of task-related licking strengthened. In trained mice, population-level representations were redundant and stable, despite dynamism of single-neuron representations. The activity of a subpopulation of neurons was consistent with touch driving licking behaviour. Our results suggest that ensembles of motor cortex neurons couple sensory input to multiple, related motor programs during learning.
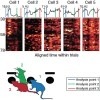
Regions within the prefrontal cortex are thought to process beliefs about the world, but little is known about the circuit dynamics underlying the formation and modification of these beliefs. Using a task that permits dissociation between the activity encoding an animal’s internal state and that encoding aspects of behavior, we found that transient increases in the volatility of activity in the rat medial prefrontal cortex accompany periods when an animal’s belief is modified after an environmental change. Activity across the majority of sampled neurons underwent marked, abrupt, and coordinated changes when prior belief was abandoned in favor of exploration of alternative strategies. These dynamics reflect network switches to a state of instability, which diminishes over the period of exploration as new stable representations are formed.
Genetically encoded calcium indicators (GECIs), together with modern microscopy, allow repeated activity measurement, in real time and with cellular resolution, of defined cellular populations. Recent efforts in protein engineering have yielded several high-quality GECIs that facilitate new applications in neuroscience. Here, we summarize recent progress in GECI design, optimization, and characterization, and provide guidelines for selecting the appropriate GECI for a given biological application. We focus on the unique challenges associated with imaging in behaving animals.
Mice lacking leptin receptors are grossly obese and diabetic, in part due to dysfunction in brain circuits important for energy homeostasis. Transplantation of leptin receptor-expressing hypothalamic progenitor neurons into the brains of leptin receptor deficient mice led to integration into neural circuits, reduced obesity, and normalized circulating glucose levels.
Our brains are capable of remarkably stable stimulus representations despite time-varying neural activity. For instance, during delay periods in working memory tasks, while stimuli are represented in working memory, neurons in the prefrontal cortex, thought to support the memory representation, exhibit time-varying neuronal activity. Since neuronal activity encodes the stimulus, its time-varying dynamics appears to be paradoxical and incompatible with stable network stimulus representations. Indeed, this finding raises a fundamental question: can stable representations only be encoded with stable neural activity, or, its corollary, is every change in activity a sign of change in stimulus representation?
We explore the hypothesis that the neuronal spike generation mechanism is an analog-to-digital converter, which rectifies low-pass filtered summed synaptic currents and encodes them into spike trains linearly decodable in post-synaptic neurons. To digitally encode an analog current waveform, the sampling rate of the spike generation mechanism must exceed its Nyquist rate. Such oversampling is consistent with the experimental observation that the precision of the spike-generation mechanism is an order of magnitude greater than the cut-off frequency of dendritic low-pass filtering. To achieve additional reduction in the error of analog-to-digital conversion, electrical engineers rely on noise-shaping. If noise-shaping were used in neurons, it would introduce correlations in spike timing to reduce low-frequency (up to Nyquist) transmission error at the cost of high-frequency one (from Nyquist to sampling rate). Using experimental data from three different classes of neurons, we demonstrate that biological neurons utilize noise-shaping. We also argue that rectification by the spike-generation mechanism may improve energy efficiency and carry out de-noising. Finally, the zoo of ion channels in neurons may be viewed as a set of predictors, various subsets of which are activated depending on the statistics of the input current.