Filter
Associated Lab
- Ahrens Lab (42) Apply Ahrens Lab filter
- Aso Lab (39) Apply Aso Lab filter
- Baker Lab (19) Apply Baker Lab filter
- Betzig Lab (98) Apply Betzig Lab filter
- Beyene Lab (4) Apply Beyene Lab filter
- Bock Lab (14) Apply Bock Lab filter
- Branson Lab (45) Apply Branson Lab filter
- Card Lab (32) Apply Card Lab filter
- Cardona Lab (44) Apply Cardona Lab filter
- Chklovskii Lab (10) Apply Chklovskii Lab filter
- Clapham Lab (10) Apply Clapham Lab filter
- Cui Lab (19) Apply Cui Lab filter
- Darshan Lab (8) Apply Darshan Lab filter
- Dickson Lab (32) Apply Dickson Lab filter
- Druckmann Lab (21) Apply Druckmann Lab filter
- Dudman Lab (34) Apply Dudman Lab filter
- Eddy/Rivas Lab (30) Apply Eddy/Rivas Lab filter
- Egnor Lab (4) Apply Egnor Lab filter
- Espinosa Medina Lab (12) Apply Espinosa Medina Lab filter
- Feliciano Lab (6) Apply Feliciano Lab filter
- Fetter Lab (31) Apply Fetter Lab filter
- Fitzgerald Lab (15) Apply Fitzgerald Lab filter
- Freeman Lab (15) Apply Freeman Lab filter
- Funke Lab (34) Apply Funke Lab filter
- Gonen Lab (59) Apply Gonen Lab filter
- Grigorieff Lab (34) Apply Grigorieff Lab filter
- Harris Lab (48) Apply Harris Lab filter
- Heberlein Lab (13) Apply Heberlein Lab filter
- Hermundstad Lab (17) Apply Hermundstad Lab filter
- Hess Lab (65) Apply Hess Lab filter
- Ilanges Lab (1) Apply Ilanges Lab filter
- Jayaraman Lab (39) Apply Jayaraman Lab filter
- Ji Lab (33) Apply Ji Lab filter
- Johnson Lab (1) Apply Johnson Lab filter
- Karpova Lab (13) Apply Karpova Lab filter
- Keleman Lab (8) Apply Keleman Lab filter
- Keller Lab (60) Apply Keller Lab filter
- Lavis Lab (120) Apply Lavis Lab filter
- Lee (Albert) Lab (29) Apply Lee (Albert) Lab filter
- Leonardo Lab (19) Apply Leonardo Lab filter
- Li Lab (1) Apply Li Lab filter
- Lippincott-Schwartz Lab (85) Apply Lippincott-Schwartz Lab filter
- Liu (Zhe) Lab (51) Apply Liu (Zhe) Lab filter
- Looger Lab (136) Apply Looger Lab filter
- Magee Lab (31) Apply Magee Lab filter
- Menon Lab (12) Apply Menon Lab filter
- Murphy Lab (6) Apply Murphy Lab filter
- O'Shea Lab (3) Apply O'Shea Lab filter
- Otopalik Lab (1) Apply Otopalik Lab filter
- Pachitariu Lab (28) Apply Pachitariu Lab filter
- Pastalkova Lab (5) Apply Pastalkova Lab filter
- Pavlopoulos Lab (7) Apply Pavlopoulos Lab filter
- Pedram Lab (2) Apply Pedram Lab filter
- Podgorski Lab (16) Apply Podgorski Lab filter
- Reiser Lab (43) Apply Reiser Lab filter
- Riddiford Lab (20) Apply Riddiford Lab filter
- Romani Lab (28) Apply Romani Lab filter
- Rubin Lab (101) Apply Rubin Lab filter
- Saalfeld Lab (41) Apply Saalfeld Lab filter
- Satou Lab (1) Apply Satou Lab filter
- Scheffer Lab (36) Apply Scheffer Lab filter
- Schreiter Lab (44) Apply Schreiter Lab filter
- Shroff Lab (21) Apply Shroff Lab filter
- Simpson Lab (18) Apply Simpson Lab filter
- Singer Lab (37) Apply Singer Lab filter
- Spruston Lab (55) Apply Spruston Lab filter
- Stern Lab (68) Apply Stern Lab filter
- Sternson Lab (47) Apply Sternson Lab filter
- Stringer Lab (23) Apply Stringer Lab filter
- Svoboda Lab (131) Apply Svoboda Lab filter
- Tebo Lab (7) Apply Tebo Lab filter
- Tervo Lab (9) Apply Tervo Lab filter
- Tillberg Lab (13) Apply Tillberg Lab filter
- Tjian Lab (17) Apply Tjian Lab filter
- Truman Lab (58) Apply Truman Lab filter
- Turaga Lab (34) Apply Turaga Lab filter
- Turner Lab (24) Apply Turner Lab filter
- Vale Lab (6) Apply Vale Lab filter
- Voigts Lab (1) Apply Voigts Lab filter
- Wang (Meng) Lab (7) Apply Wang (Meng) Lab filter
- Wang (Shaohe) Lab (2) Apply Wang (Shaohe) Lab filter
- Wu Lab (8) Apply Wu Lab filter
- Zlatic Lab (26) Apply Zlatic Lab filter
- Zuker Lab (5) Apply Zuker Lab filter
Associated Project Team
- CellMap (1) Apply CellMap filter
- COSEM (3) Apply COSEM filter
- Fly Descending Interneuron (10) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (14) Apply Fly Functional Connectome filter
- Fly Olympiad (5) Apply Fly Olympiad filter
- FlyEM (50) Apply FlyEM filter
- FlyLight (46) Apply FlyLight filter
- GENIE (40) Apply GENIE filter
- Integrative Imaging (1) Apply Integrative Imaging filter
- Larval Olympiad (2) Apply Larval Olympiad filter
- MouseLight (16) Apply MouseLight filter
- NeuroSeq (1) Apply NeuroSeq filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (22) Apply Tool Translation Team (T3) filter
- Transcription Imaging (45) Apply Transcription Imaging filter
Associated Support Team
- Anatomy and Histology (18) Apply Anatomy and Histology filter
- Cryo-Electron Microscopy (33) Apply Cryo-Electron Microscopy filter
- Electron Microscopy (11) Apply Electron Microscopy filter
- Fly Facility (39) Apply Fly Facility filter
- Gene Targeting and Transgenics (10) Apply Gene Targeting and Transgenics filter
- Integrative Imaging (10) Apply Integrative Imaging filter
- Janelia Experimental Technology (35) Apply Janelia Experimental Technology filter
- Management Team (1) Apply Management Team filter
- Molecular Genomics (15) Apply Molecular Genomics filter
- Primary & iPS Cell Culture (13) Apply Primary & iPS Cell Culture filter
- Project Technical Resources (32) Apply Project Technical Resources filter
- Quantitative Genomics (18) Apply Quantitative Genomics filter
- Scientific Computing Software (56) Apply Scientific Computing Software filter
- Scientific Computing Systems (6) Apply Scientific Computing Systems filter
- Viral Tools (14) Apply Viral Tools filter
- Vivarium (6) Apply Vivarium filter
Publication Date
- 2024 (102) Apply 2024 filter
- 2023 (177) Apply 2023 filter
- 2022 (166) Apply 2022 filter
- 2021 (174) Apply 2021 filter
- 2020 (178) Apply 2020 filter
- 2019 (177) Apply 2019 filter
- 2018 (206) Apply 2018 filter
- 2017 (186) Apply 2017 filter
- 2016 (191) Apply 2016 filter
- 2015 (195) Apply 2015 filter
- 2014 (190) Apply 2014 filter
- 2013 (136) Apply 2013 filter
- 2012 (112) Apply 2012 filter
- 2011 (98) Apply 2011 filter
- 2010 (61) Apply 2010 filter
- 2009 (56) Apply 2009 filter
- 2008 (40) Apply 2008 filter
- 2007 (21) Apply 2007 filter
- 2006 (3) Apply 2006 filter
2469 Janelia Publications
Showing 2431-2440 of 2469 resultsGenetically-encoded calcium indicators (GECIs) hold the promise of monitoring [Ca(2+)] in selected populations of neurons and in specific cellular compartments. Relating GECI fluorescence to neuronal activity requires quantitative characterization. We have characterized a promising new genetically-encoded calcium indicator-GCaMP2-in mammalian pyramidal neurons. Fluorescence changes in response to single action potentials (17+/-10% DeltaF/F [mean+/-SD]) could be detected in some, but not all, neurons. Trains of high-frequency action potentials yielded robust responses (302+/-50% for trains of 40 action potentials at 83 Hz). Responses were similar in acute brain slices from in utero electroporated mice, indicating that long-term expression did not interfere with GCaMP2 function. Membrane-targeted versions of GCaMP2 did not yield larger signals than their non-targeted counterparts. We further targeted GCaMP2 to dendritic spines to monitor Ca(2+) accumulations evoked by activation of synaptic NMDA receptors. We observed robust DeltaF/F responses (range: 37%-264%) to single spine uncaging stimuli that were correlated with NMDA receptor currents measured through a somatic patch pipette. One major drawback of GCaMP2 was its low baseline fluorescence. Our results show that GCaMP2 is improved from the previous versions of GCaMP and may be suited to detect bursts of high-frequency action potentials and synaptic currents in vivo.
Understanding the principles of information processing in neural circuits requires systematic characterization of the participating cell types and their connections, and the ability to measure and perturb their activity. Genetic approaches promise to bring experimental access to complex neural systems, including genetic stalwarts such as the fly and mouse, but also to nongenetic systems such as primates. Together with anatomical and physiological methods, cell-type-specific expression of protein markers and sensors and transducers will be critical to construct circuit diagrams and to measure the activity of genetically defined neurons. Inactivation and activation of genetically defined cell types will establish causal relationships between activity in specific groups of neurons, circuit function, and animal behavior. Genetic analysis thus promises to reveal the logic of the neural circuits in complex brains that guide behaviors. Here we review progress in the genetic analysis of neural circuits and discuss directions for future research and development.
Pyramidal neurons are characterized by their distinct apical and basal dendritic trees and the pyramidal shape of their soma. They are found in several regions of the CNS and, although the reasons for their abundance remain unclear, functional studies--especially of CA1 hippocampal and layer V neocortical pyramidal neurons--have offered insights into the functions of their unique cellular architecture. Pyramidal neurons are not all identical, but some shared functional principles can be identified. In particular, the existence of dendritic domains with distinct synaptic inputs, excitability, modulation and plasticity appears to be a common feature that allows synapses throughout the dendritic tree to contribute to action-potential generation. These properties support a variety of coincidence-detection mechanisms, which are likely to be crucial for synaptic integration and plasticity.
The identification of synaptic partners is challenging in dense nerve bundles, where many processes occupy regions beneath the resolution of conventional light microscopy. To address this difficulty, we have developed GRASP, a system to label membrane contacts and synapses between two cells in living animals. Two complementary fragments of GFP are expressed on different cells, tethered to extracellular domains of transmembrane carrier proteins. When the complementary GFP fragments are fused to ubiquitous transmembrane proteins, GFP fluorescence appears uniformly along membrane contacts between the two cells. When one or both GFP fragments are fused to synaptic transmembrane proteins, GFP fluorescence is tightly localized to synapses. GRASP marks known synaptic contacts in C. elegans, correctly identifies changes in mutants with altered synaptic specificity, and can uncover new information about synaptic locations as confirmed by electron microscopy. GRASP may prove particularly useful for defining connectivity in complex nervous systems.
We performed patch-clamp recordings from morphologically identified and anatomically mapped pyramidal neurons of the ventral hippocampus to test the hypothesis that bursting neurons are distributed on a gradient from the CA2/CA1 border (proximal) through the subiculum (distal), with more bursting observed at distal locations. We find that the well-defined morphological boundaries between the hippocampal subregions CA1 and subiculum do not correspond to abrupt changes in electrophysiological properties. Rather, we observed that the percentage of bursting neurons is linearly correlated with position in the proximal-distal axis across the CA1 and the subiculum, the percentages of bursting neurons being 10% near the CA1-CA2 border, 24% at the CA1-subiculum border, and higher than 50% in the distal subiculum. The distribution of bursting neurons was paralleled by a gradient in afterdepolarization (ADP) amplitude. We also tested the hypothesis that there was an association between bursting and two previously described morphologically distinct groups of pyramidal neurons (twin and single apical dendrites) in the CA1 region. We found no difference in output mode between single and twin apical dendrite morphologies, which was consistent with the observation that the two morphologies were equally distributed across the transverse axis of the CA1 region. Taken together with the known organization of connections from CA3 to CA1 and CA1 to subiculum, our results indicate that bursting neurons are most likely to be connected to regular spiking neurons and vice versa.
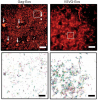
We combined photoactivated localization microscopy (PALM) with live-cell single-particle tracking to create a new method termed sptPALM. We created spatially resolved maps of single-molecule motions by imaging the membrane proteins Gag and VSVG, and obtained several orders of magnitude more trajectories per cell than traditional single-particle tracking enables. By probing distinct subsets of molecules, sptPALM can provide insight into the origins of spatial and temporal heterogeneities in membranes.
Commentary: As a stepping stone to true live cell PALM (see above), our collaborator Jennifer Lippincott-Schwartz suggested using the sparse photoactivation principle of PALM to track the nanoscale motion of thousands of individual molecules within a single living cell. Termed single particle tracking PALM (sptPALM), Jennifer’s postdocs Suliana Manley and Jen Gillette used the method in our PALM rig to create spatially resolved maps of diffusion rates in the plasma membrane of live cells. sptPALM is a powerful tool to study the active cytoskeletal or passive diffusional transport of individual molecules with far more measurements per cell than is possible without sparse photoactivation.
Pulsed lasers are key elements in nonlinear bioimaging techniques such as two-photon fluorescence excitation (TPE) microscopy. Typically, however, only a percent or less of the laser power available can be delivered to the sample before photoinduced damage becomes excessive. Here we describe a passive pulse splitter that converts each laser pulse into a fixed number of sub-pulses of equal energy. We applied the splitter to TPE imaging of fixed mouse brain slices labeled with GFP and show that, in different power regimes, the splitter can be used either to increase the signal rate more than 100-fold or to reduce the rate of photobleaching by over fourfold. In living specimens, the gains were even greater: a ninefold reduction in photobleaching during in vivo imaging of Caenorhabditis elegans larvae, and a six- to 20-fold decrease in the rate of photodamage during calcium imaging of rat hippocampal brain slices.

Pulsed lasers are key elements in nonlinear bioimaging techniques such as two-photon fluorescence excitation (TPE) microscopy. Typically, however, only a percent or less of the laser power available can be delivered to the sample before photoinduced damage becomes excessive. Here we describe a passive pulse splitter that converts each laser pulse into a fixed number of sub-pulses of equal energy. We applied the splitter to TPE imaging of fixed mouse brain slices labeled with GFP and show that, in different power regimes, the splitter can be used either to increase the signal rate more than 100-fold or to reduce the rate of photobleaching by over fourfold. In living specimens, the gains were even greater: a ninefold reduction in photobleaching during in vivo imaging of Caenorhabditis elegans larvae, and a six- to 20-fold decrease in the rate of photodamage during calcium imaging of rat hippocampal brain slices.
Commentary: Na Ji came to me early in her postdoc with an idea to reduce photodamage in nonlinear microscopy by splitting the pulses from an ultrafast laser into multiple subpulses of reduced energy. In six weeks, we constructed a prototype pulse splitter and obtained initial results confirming the validity of her vision. Further experiments with Jeff Magee demonstrated that the splitter could be used to increase imaging speed or reduce photodamage in two photon microscopy by one to two orders of magnitude. This project is a great example of how quickly one can react and exploit new ideas in the Janelia environment.
MOTIVATION: Caenorhabditis elegans, a roundworm found in soil, is a widely studied model organism with about 1000 cells in the adult. Producing high-resolution fluorescence images of C.elegans to reveal biological insights is becoming routine, motivating the development of advanced computational tools for analyzing the resulting image stacks. For example, worm bodies usually curve significantly in images. Thus one must ’straighten’ the worms if they are to be compared under a canonical coordinate system. RESULTS: We develop a worm straightening algorithm (WSA) that restacks cutting planes orthogonal to a ’backbone’ that models the anterior-posterior axis of the worm. We formulate the backbone as a parametric cubic spline defined by a series of control points. We develop two methods for automatically determining the locations of the control points. Our experimental methods show that our approaches effectively straighten both 2D and 3D worm images.