Filter
Associated Lab
- Aguilera Castrejon Lab (1) Apply Aguilera Castrejon Lab filter
- Ahrens Lab (53) Apply Ahrens Lab filter
- Aso Lab (40) Apply Aso Lab filter
- Baker Lab (19) Apply Baker Lab filter
- Betzig Lab (101) Apply Betzig Lab filter
- Beyene Lab (8) Apply Beyene Lab filter
- Bock Lab (14) Apply Bock Lab filter
- Branson Lab (50) Apply Branson Lab filter
- Card Lab (36) Apply Card Lab filter
- Cardona Lab (45) Apply Cardona Lab filter
- Chklovskii Lab (10) Apply Chklovskii Lab filter
- Clapham Lab (14) Apply Clapham Lab filter
- Cui Lab (19) Apply Cui Lab filter
- Darshan Lab (8) Apply Darshan Lab filter
- Dickson Lab (32) Apply Dickson Lab filter
- Druckmann Lab (21) Apply Druckmann Lab filter
- Dudman Lab (38) Apply Dudman Lab filter
- Eddy/Rivas Lab (30) Apply Eddy/Rivas Lab filter
- Egnor Lab (4) Apply Egnor Lab filter
- Espinosa Medina Lab (15) Apply Espinosa Medina Lab filter
- Feliciano Lab (7) Apply Feliciano Lab filter
- Fetter Lab (31) Apply Fetter Lab filter
- Fitzgerald Lab (16) Apply Fitzgerald Lab filter
- Freeman Lab (15) Apply Freeman Lab filter
- Funke Lab (38) Apply Funke Lab filter
- Gonen Lab (59) Apply Gonen Lab filter
- Grigorieff Lab (34) Apply Grigorieff Lab filter
- Harris Lab (53) Apply Harris Lab filter
- Heberlein Lab (13) Apply Heberlein Lab filter
- Hermundstad Lab (23) Apply Hermundstad Lab filter
- Hess Lab (74) Apply Hess Lab filter
- Ilanges Lab (2) Apply Ilanges Lab filter
- Jayaraman Lab (42) Apply Jayaraman Lab filter
- Ji Lab (33) Apply Ji Lab filter
- Johnson Lab (1) Apply Johnson Lab filter
- Karpova Lab (13) Apply Karpova Lab filter
- Keleman Lab (8) Apply Keleman Lab filter
- Keller Lab (61) Apply Keller Lab filter
- Koay Lab (2) Apply Koay Lab filter
- Lavis Lab (137) Apply Lavis Lab filter
- Lee (Albert) Lab (29) Apply Lee (Albert) Lab filter
- Leonardo Lab (19) Apply Leonardo Lab filter
- Li Lab (4) Apply Li Lab filter
- Lippincott-Schwartz Lab (97) Apply Lippincott-Schwartz Lab filter
- Liu (Yin) Lab (1) Apply Liu (Yin) Lab filter
- Liu (Zhe) Lab (58) Apply Liu (Zhe) Lab filter
- Looger Lab (137) Apply Looger Lab filter
- Magee Lab (31) Apply Magee Lab filter
- Menon Lab (12) Apply Menon Lab filter
- Murphy Lab (6) Apply Murphy Lab filter
- O'Shea Lab (6) Apply O'Shea Lab filter
- Otopalik Lab (1) Apply Otopalik Lab filter
- Pachitariu Lab (36) Apply Pachitariu Lab filter
- Pastalkova Lab (5) Apply Pastalkova Lab filter
- Pavlopoulos Lab (7) Apply Pavlopoulos Lab filter
- Pedram Lab (4) Apply Pedram Lab filter
- Podgorski Lab (16) Apply Podgorski Lab filter
- Reiser Lab (45) Apply Reiser Lab filter
- Riddiford Lab (20) Apply Riddiford Lab filter
- Romani Lab (31) Apply Romani Lab filter
- Rubin Lab (105) Apply Rubin Lab filter
- Saalfeld Lab (46) Apply Saalfeld Lab filter
- Satou Lab (1) Apply Satou Lab filter
- Scheffer Lab (36) Apply Scheffer Lab filter
- Schreiter Lab (51) Apply Schreiter Lab filter
- Sgro Lab (1) Apply Sgro Lab filter
- Shroff Lab (31) Apply Shroff Lab filter
- Simpson Lab (18) Apply Simpson Lab filter
- Singer Lab (37) Apply Singer Lab filter
- Spruston Lab (57) Apply Spruston Lab filter
- Stern Lab (73) Apply Stern Lab filter
- Sternson Lab (47) Apply Sternson Lab filter
- Stringer Lab (33) Apply Stringer Lab filter
- Svoboda Lab (131) Apply Svoboda Lab filter
- Tebo Lab (9) Apply Tebo Lab filter
- Tervo Lab (9) Apply Tervo Lab filter
- Tillberg Lab (18) Apply Tillberg Lab filter
- Tjian Lab (17) Apply Tjian Lab filter
- Truman Lab (58) Apply Truman Lab filter
- Turaga Lab (39) Apply Turaga Lab filter
- Turner Lab (27) Apply Turner Lab filter
- Vale Lab (7) Apply Vale Lab filter
- Voigts Lab (3) Apply Voigts Lab filter
- Wang (Meng) Lab (21) Apply Wang (Meng) Lab filter
- Wang (Shaohe) Lab (6) Apply Wang (Shaohe) Lab filter
- Wu Lab (8) Apply Wu Lab filter
- Zlatic Lab (26) Apply Zlatic Lab filter
- Zuker Lab (5) Apply Zuker Lab filter
Associated Project Team
- CellMap (12) Apply CellMap filter
- COSEM (3) Apply COSEM filter
- FIB-SEM Technology (3) Apply FIB-SEM Technology filter
- Fly Descending Interneuron (11) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (14) Apply Fly Functional Connectome filter
- Fly Olympiad (5) Apply Fly Olympiad filter
- FlyEM (53) Apply FlyEM filter
- FlyLight (49) Apply FlyLight filter
- GENIE (46) Apply GENIE filter
- Integrative Imaging (4) Apply Integrative Imaging filter
- Larval Olympiad (2) Apply Larval Olympiad filter
- MouseLight (18) Apply MouseLight filter
- NeuroSeq (1) Apply NeuroSeq filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (26) Apply Tool Translation Team (T3) filter
- Transcription Imaging (45) Apply Transcription Imaging filter
Associated Support Team
- Project Pipeline Support (5) Apply Project Pipeline Support filter
- Anatomy and Histology (18) Apply Anatomy and Histology filter
- Cryo-Electron Microscopy (37) Apply Cryo-Electron Microscopy filter
- Electron Microscopy (16) Apply Electron Microscopy filter
- Gene Targeting and Transgenics (11) Apply Gene Targeting and Transgenics filter
- Integrative Imaging (17) Apply Integrative Imaging filter
- Invertebrate Shared Resource (40) Apply Invertebrate Shared Resource filter
- Janelia Experimental Technology (37) Apply Janelia Experimental Technology filter
- Management Team (1) Apply Management Team filter
- Molecular Genomics (15) Apply Molecular Genomics filter
- Primary & iPS Cell Culture (14) Apply Primary & iPS Cell Culture filter
- Project Technical Resources (50) Apply Project Technical Resources filter
- Quantitative Genomics (19) Apply Quantitative Genomics filter
- Scientific Computing Software (92) Apply Scientific Computing Software filter
- Scientific Computing Systems (7) Apply Scientific Computing Systems filter
- Viral Tools (14) Apply Viral Tools filter
- Vivarium (7) Apply Vivarium filter
Publication Date
- 2025 (130) Apply 2025 filter
- 2024 (215) Apply 2024 filter
- 2023 (159) Apply 2023 filter
- 2022 (167) Apply 2022 filter
- 2021 (175) Apply 2021 filter
- 2020 (177) Apply 2020 filter
- 2019 (177) Apply 2019 filter
- 2018 (206) Apply 2018 filter
- 2017 (186) Apply 2017 filter
- 2016 (191) Apply 2016 filter
- 2015 (195) Apply 2015 filter
- 2014 (190) Apply 2014 filter
- 2013 (136) Apply 2013 filter
- 2012 (112) Apply 2012 filter
- 2011 (98) Apply 2011 filter
- 2010 (61) Apply 2010 filter
- 2009 (56) Apply 2009 filter
- 2008 (40) Apply 2008 filter
- 2007 (21) Apply 2007 filter
- 2006 (3) Apply 2006 filter
2695 Janelia Publications
Showing 2571-2580 of 2695 resultsIt is well known that light-sheet illumination can enable optically sectioned wide-field imaging of macroscopic samples. However, the optical sectioning capacity of a light-sheet macroscope is undermined by sample-induced scattering or aberrations that broaden the thickness of the sheet illumination. We present a technique to enhance the optical sectioning capacity of a scanning light-sheet microscope by out-of-focus background rejection. The technique, called HiLo microscopy, makes use of two images sequentially acquired with uniform and structured sheet illumination. An optically sectioned image is then synthesized by fusing high and low spatial frequency information from both images. The benefits of combining light-sheet macroscopy and HiLo background rejection are demonstrated in optically cleared whole mouse brain samples, using both green fluorescent protein (GFP)-fluorescence and dark-field scattered light contrast.
BACKGROUND: Previous studies suggest that mechanical feedback could coordinate morphogenetic events in embryos. Furthermore, embryonic tissues have complex structure and composition and undergo large deformations during morphogenesis. Hence we expect highly non-linear and loading-rate dependent tissue mechanical properties in embryos. METHODOLOGY/PRINCIPAL FINDINGS: We used micro-aspiration to test whether a simple linear viscoelastic model was sufficient to describe the mechanical behavior of gastrula stage Xenopus laevis embryonic tissue in vivo. We tested whether these embryonic tissues change their mechanical properties in response to mechanical stimuli but found no evidence of changes in the viscoelastic properties of the tissue in response to stress or stress application rate. We used this model to test hypotheses about the pattern of force generation during electrically induced tissue contractions. The dependence of contractions on suction pressure was most consistent with apical tension, and was inconsistent with isotropic contraction. Finally, stiffer clutches generated stronger contractions, suggesting that force generation and stiffness may be coupled in the embryo. CONCLUSIONS/SIGNIFICANCE: The mechanical behavior of a complex, active embryonic tissue can be surprisingly well described by a simple linear viscoelastic model with power law creep compliance, even at high deformations. We found no evidence of mechanical feedback in this system. Together these results show that very simple mechanical models can be useful in describing embryo mechanics.
BACKGROUND: Conditional gene activation is an efficient strategy for studying gene function in genetically modified animals. Among the presently available gene switches, the tetracycline-regulated system has attracted considerable interest because of its unique potential for reversible and adjustable gene regulation. RESULTS: To investigate whether the ubiquitously expressed Gt(ROSA)26Sor locus enables uniform DOX-controlled gene expression, we inserted the improved tetracycline-regulated transcription activator iM2 together with an iM2 dependent GFP gene into the Gt(ROSA)26Sor locus, using gene targeting to generate ROSA26-iM2-GFP (R26t1Δ) mice. Despite the presence of ROSA26 promoter driven iM2, R26t1Δ mice showed very sparse DOX-activated expression of different iM2-responsive reporter genes in the brain, mosaic expression in peripheral tissues and more prominent expression in erythroid, myeloid and lymphoid lineages, in hematopoietic stem and progenitor cells and in olfactory neurons. CONCLUSIONS: The finding that gene regulation by the DOX-activated transcriptional factor iM2 in the Gt(ROSA)26Sor locus has its limitations is of importance for future experimental strategies involving transgene activation from the endogenous ROSA26 promoter. Furthermore, our ROSA26-iM2 knock-in mouse model (R26t1Δ) represents a useful tool for implementing gene function in vivo especially under circumstances requiring the side-by-side comparison of gene manipulated and wild type cells. Since the ROSA26-iM2 mouse allows mosaic gene activation in peripheral tissues and haematopoietic cells, this model will be very useful for uncovering previously unknown or unsuspected phenotypes.
Pfam is a widely used database of protein families and domains. This article describes a set of major updates that we have implemented in the latest release (version 24.0). The most important change is that we now use HMMER3, the latest version of the popular profile hidden Markov model package. This software is approximately 100 times faster than HMMER2 and is more sensitive due to the routine use of the forward algorithm. The move to HMMER3 has necessitated numerous changes to Pfam that are described in detail. Pfam release 24.0 contains 11,912 families, of which a large number have been significantly updated during the past two years. Pfam is available via servers in the UK (http://pfam.sanger.ac.uk/), the USA (http://pfam.janelia.org/) and Sweden (http://pfam.sbc.su.se/).
Animals generate diverse motor behaviors, yet how the same motor neurons (MNs) generate two distinct or antagonistic behaviors remains an open question. Here we characterize larval muscle activity patterns and premotor/motor circuits to understand how they generate forward and backward locomotion. We show that all body wall MNs are activated during both behaviors, but a subset of MNs change recruitment timing for each behavior. We used TEM to reconstruct a full segment of all 60 MNs and 236 premotor neurons (PMNs), including differentially-recruited MNs. Analysis of this comprehensive connectome identified PMN-MN 'labeled line' connectivity; PMN-MN combinatorial connectivity; asymmetric neuronal morphology; and PMN-MN circuit motifs that could all contribute to generating distinct behaviors. We generated a recurrent network model that reproduced the observed behaviors, and used functional optogenetics to validate selected model predictions. This PMN-MN connectome will provide a foundation for analyzing the full suite of larval behaviors.
Nuclear receptors (NRs) comprise a family of ligand-regulated transcription factors that control diverse critical biological processes including various aspects of brain development. Eighteen NR genes exist in the Drosophila genome. To explore their roles in brain development, we knocked down individual NRs through the development of the mushroom bodies (MBs) by targeted RNAi. Besides recapitulating the known MB phenotypes for three NRs, we found that unfulfilled (unf), an ortholog of human photoreceptor specific nuclear receptor (PNR), regulates axonal morphogenesis and neuronal subtype identity. The adult MBs develop through remodeling of gamma neurons plus de-novo elaboration of both alpha’/beta’ and alpha/beta neurons. Notably, unf is largely dispensable for the initial elaboration of gamma neurons, but plays an essential role in their re-extension of axons after pruning during early metamorphosis. The subsequently derived MB neuron types also require unf for extension of axons beyond the terminus of the pruned bundle. Tracing single axons revealed misrouting rather than simple truncation. Further, silencing unf in single-cell clones elicited misguidance of axons in otherwise unperturbed MBs. Such axon guidance defects may occur as MB neurons partially lose their subtype identity, as evidenced by suppression of various MB subtype markers in unf knockdown MBs. In sum, unf governs axonal morphogenesis of multiple MB neuron types, possibly through regulating neuronal subtype identity.
The microtubule cytoskeleton has proven to be an effective target for cancer therapeutics. One class of drugs, known as microtubule stabilizing agents (MSAs), binds to microtubule polymers and stabilizes them against depolymerization. The prototype of this group of drugs, Taxol, is an effective chemotherapeutic agent used extensively in the treatment of human ovarian, breast, and lung carcinomas. Although electron crystallography and photoaffinity labeling experiments determined that the binding site for Taxol is in a hydrophobic pocket in beta-tubulin, little was known about the effects of this drug on the conformation of the entire microtubule. A recent study from our laboratory utilizing hydrogen-deuterium exchange (HDX) in concert with various mass spectrometry (MS) techniques has provided new information on the structure of microtubules upon Taxol binding. In the current study we apply this technique to determine the binding mode and the conformational effects on chicken erythrocyte tubulin (CET) of another MSA, discodermolide, whose synthetic analogues may have potential use in the clinic. We confirmed that, like Taxol, discodermolide binds to the taxane binding pocket in beta-tubulin. However, as opposed to Taxol, which has major interactions with the M-loop, discodermolide orients itself away from this loop and toward the N-terminal H1-S2 loop. Additionally, discodermolide stabilizes microtubules mainly via its effects on interdimer contacts, specifically on the alpha-tubulin side, and to a lesser extent on interprotofilament contacts between adjacent beta-tubulin subunits. Also, our results indicate complementary stabilizing effects of Taxol and discodermolide on the microtubules, which may explain the synergy observed between the two drugs in vivo.
The excitability of individual dendritic branches is a plastic property of neurons. We found that experience in an enriched environment increased propagation of dendritic Na(+) spikes in a subset of individual dendritic branches in rat hippocampal CA1 pyramidal neurons and that this effect was mainly mediated by localized downregulation of A-type K(+) channel function. Thus, dendritic plasticity might be used to store recent experience in individual branches of the dendritic arbor.
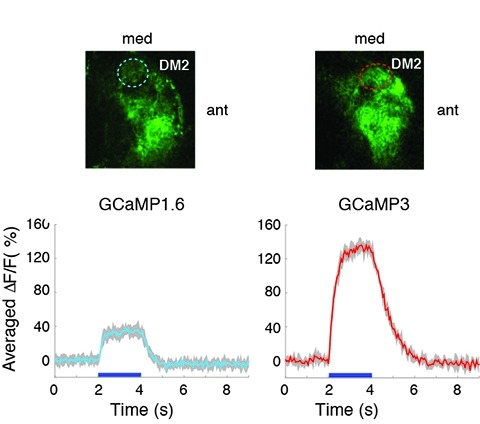
Genetically encoded calcium indicators (GECIs) can be used to image activity in defined neuronal populations. However, current GECIs produce inferior signals compared to synthetic indicators and recording electrodes, precluding detection of low firing rates. We developed a single-wavelength GCaMP2-based GECI (GCaMP3), with increased baseline fluorescence (3-fold), increased dynamic range (3-fold) and higher affinity for calcium (1.3-fold). We detected GCaMP3 fluorescence changes triggered by single action potentials in pyramidal cell dendrites, with signal-to-noise ratio and photostability substantially better than those of GCaMP2, D3cpVenus and TN-XXL. In Caenorhabditis elegans chemosensory neurons and the Drosophila melanogaster antennal lobe, sensory stimulation-evoked fluorescence responses were significantly enhanced with GCaMP3 (4-6-fold). In somatosensory and motor cortical neurons in the intact mouse, GCaMP3 detected calcium transients with amplitudes linearly dependent on action potential number. Long-term imaging in the motor cortex of behaving mice revealed large fluorescence changes in imaged neurons over months.