Filter
Associated Lab
- Ahrens Lab (5) Apply Ahrens Lab filter
- Aso Lab (3) Apply Aso Lab filter
- Betzig Lab (7) Apply Betzig Lab filter
- Bock Lab (5) Apply Bock Lab filter
- Branson Lab (3) Apply Branson Lab filter
- Card Lab (2) Apply Card Lab filter
- Cardona Lab (4) Apply Cardona Lab filter
- Clapham Lab (2) Apply Clapham Lab filter
- Darshan Lab (1) Apply Darshan Lab filter
- Dickson Lab (5) Apply Dickson Lab filter
- Druckmann Lab (3) Apply Druckmann Lab filter
- Dudman Lab (4) Apply Dudman Lab filter
- Espinosa Medina Lab (3) Apply Espinosa Medina Lab filter
- Feliciano Lab (1) Apply Feliciano Lab filter
- Fitzgerald Lab (2) Apply Fitzgerald Lab filter
- Funke Lab (1) Apply Funke Lab filter
- Gonen Lab (2) Apply Gonen Lab filter
- Grigorieff Lab (4) Apply Grigorieff Lab filter
- Harris Lab (4) Apply Harris Lab filter
- Heberlein Lab (2) Apply Heberlein Lab filter
- Hermundstad Lab (1) Apply Hermundstad Lab filter
- Hess Lab (5) Apply Hess Lab filter
- Jayaraman Lab (4) Apply Jayaraman Lab filter
- Ji Lab (1) Apply Ji Lab filter
- Johnson Lab (1) Apply Johnson Lab filter
- Keleman Lab (2) Apply Keleman Lab filter
- Keller Lab (6) Apply Keller Lab filter
- Lavis Lab (6) Apply Lavis Lab filter
- Lee (Albert) Lab (1) Apply Lee (Albert) Lab filter
- Lippincott-Schwartz Lab (12) Apply Lippincott-Schwartz Lab filter
- Liu (Zhe) Lab (7) Apply Liu (Zhe) Lab filter
- Looger Lab (15) Apply Looger Lab filter
- O'Shea Lab (1) Apply O'Shea Lab filter
- Pachitariu Lab (4) Apply Pachitariu Lab filter
- Pavlopoulos Lab (1) Apply Pavlopoulos Lab filter
- Podgorski Lab (4) Apply Podgorski Lab filter
- Reiser Lab (2) Apply Reiser Lab filter
- Romani Lab (3) Apply Romani Lab filter
- Rubin Lab (6) Apply Rubin Lab filter
- Saalfeld Lab (3) Apply Saalfeld Lab filter
- Scheffer Lab (2) Apply Scheffer Lab filter
- Schreiter Lab (4) Apply Schreiter Lab filter
- Simpson Lab (1) Apply Simpson Lab filter
- Singer Lab (4) Apply Singer Lab filter
- Spruston Lab (6) Apply Spruston Lab filter
- Stern Lab (5) Apply Stern Lab filter
- Sternson Lab (2) Apply Sternson Lab filter
- Stringer Lab (3) Apply Stringer Lab filter
- Svoboda Lab (14) Apply Svoboda Lab filter
- Tillberg Lab (2) Apply Tillberg Lab filter
- Truman Lab (4) Apply Truman Lab filter
- Turaga Lab (2) Apply Turaga Lab filter
- Turner Lab (2) Apply Turner Lab filter
- Zlatic Lab (1) Apply Zlatic Lab filter
Associated Project Team
Publication Date
- December 2019 (9) Apply December 2019 filter
- November 2019 (11) Apply November 2019 filter
- October 2019 (18) Apply October 2019 filter
- September 2019 (15) Apply September 2019 filter
- August 2019 (14) Apply August 2019 filter
- July 2019 (12) Apply July 2019 filter
- June 2019 (18) Apply June 2019 filter
- May 2019 (12) Apply May 2019 filter
- April 2019 (16) Apply April 2019 filter
- March 2019 (17) Apply March 2019 filter
- February 2019 (18) Apply February 2019 filter
- January 2019 (17) Apply January 2019 filter
- Remove 2019 filter 2019
Type of Publication
- Remove Janelia filter Janelia
177 Publications
Showing 71-80 of 177 resultsFocused Ion Beam Scanning Electron Microscopy (FIB-SEM) generates 3D datasets optimally suited for segmentation of cell ultrastructure and automated connectome tracing but is limited to small fields of view and is therefore incompatible with the new generation of ultrafast multibeam SEMs. In contrast, section-based techniques are multibeam-compatible but are limited in z-resolution making automatic segmentation of cellular ultrastructure difficult. Here we demonstrate a novel 3D electron microscopy technique, Gas Cluster Ion Beam SEM (GCIB-SEM), in which top-down, wide-area ion milling is performed on a series of thick sections, acquiring < 10 nm isotropic datasets of each which are then stitched together to span the full sectioned volume. Based on our results, incorporating GCIB-SEM into existing single beam and multibeam SEM workflows should be straightforward and should dramatically increase reliability while simultaneously improving z-resolution by a factor of 3 or more.
View Publication Page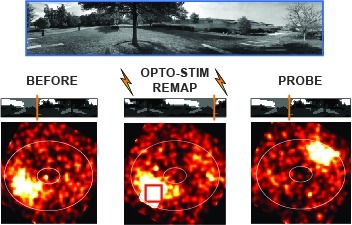
Many animals rely on an internal heading representation when navigating in varied environments. How this representation is linked to the sensory cues that define different surroundings is unclear. In the fly brain, heading is represented by 'compass' neurons that innervate a ring-shaped structure known as the ellipsoid body. Each compass neuron receives inputs from 'ring' neurons that are selective for particular visual features; this combination provides an ideal substrate for the extraction of directional information from a visual scene. Here we combine two-photon calcium imaging and optogenetics in tethered flying flies with circuit modelling, and show how the correlated activity of compass and visual neurons drives plasticity, which flexibly transforms two-dimensional visual cues into a stable heading representation. We also describe how this plasticity enables the fly to convert a partial heading representation, established from orienting within part of a novel setting, into a complete heading representation. Our results provide mechanistic insight into the memory-related computations that are essential for flexible navigation in varied surroundings.
Different memory components are forgotten through distinct molecular mechanisms. In , the activation of 2 Rho GTPases (Rac1 and Cdc42), respectively, underlies the forgetting of an early labile memory (anesthesia-sensitive memory, ASM) and a form of consolidated memory (anesthesia-resistant memory, ARM). Here, we dissected the molecular mechanisms that tie Rac1 and Cdc42 to the different types of memory forgetting. We found that 2 WASP family proteins, SCAR/WAVE and WASp, act downstream of Rac1 and Cdc42 separately to regulate ASM and ARM forgetting in mushroom body neurons. Arp2/3 complex, which organizes branched actin polymerization, is a canonical downstream effector of WASP family proteins. However, we found that Arp2/3 complex is required in Cdc42/WASp-mediated ARM forgetting but not in Rac1/SCAR-mediated ASM forgetting. Instead, we identified that Rac1/SCAR may function with formin Diaphanous (Dia), a nucleator that facilitates linear actin polymerization, in ASM forgetting. The present study, complementing the previously identified Rac1/cofilin pathway that regulates actin depolymerization, suggests that Rho GTPases regulate forgetting by recruiting both actin polymerization and depolymerization pathways. Moreover, Rac1 and Cdc42 may regulate different types of memory forgetting by tapping into different actin polymerization mechanisms.
Autosomal recessive (AR) gene defects are the leading genetic cause of intellectual disability (ID) in countries with frequent parental consanguinity, which account for about 1/7th of the world population. Yet, compared to autosomal dominant de novo mutations, which are the predominant cause of ID in Western countries, the identification of AR-ID genes has lagged behind. Here, we report on whole exome and whole genome sequencing in 404 consanguineous predominantly Iranian families with two or more affected offspring. In 219 of these, we found likely causative variants, involving 77 known and 77 novel AR-ID (candidate) genes, 21 X-linked genes, as well as 9 genes previously implicated in diseases other than ID. This study, the largest of its kind published to date, illustrates that high-throughput DNA sequencing in consanguineous families is a superior strategy for elucidating the thousands of hitherto unknown gene defects underlying AR-ID, and it sheds light on their prevalence.
When a behavior repeatedly fails to achieve its goal, animals often give up and become passive, which can be strategic for preserving energy or regrouping between attempts. It is unknown how the brain identifies behavioral failures and mediates this behavioral-state switch. In larval zebrafish swimming in virtual reality, visual feedback can be withheld so that swim attempts fail to trigger expected visual flow. After tens of seconds of such motor futility, animals became passive for similar durations. Whole-brain calcium imaging revealed noradrenergic neurons that responded specifically to failed swim attempts and radial astrocytes whose calcium levels accumulated with increasing numbers of failed attempts. Using cell ablation and optogenetic or chemogenetic activation, we found that noradrenergic neurons progressively activated brainstem radial astrocytes, which then suppressed swimming. Thus, radial astrocytes perform a computation critical for behavior: they accumulate evidence that current actions are ineffective and consequently drive changes in behavioral states.
The mechanistic operation of brain regions is often interpreted by partitioning constituent neurons into 'cell types'. Historically, such cell types were broadly defined by their correspondence to gross features of the nervous system (such as cytoarchitecture). Modern-day neuroscientific techniques, enabling a more nuanced examination of neuronal properties, have illustrated a wealth of heterogeneity within these classical cell types. Here, we review the extent of this within-cell-type heterogeneity in one of the simplest cortical regions of the mammalian brain, the rodent hippocampus. We focus on the mounting evidence that the classical CA3, CA1 and subiculum pyramidal cell types all exhibit prominent and spatially patterned within-cell-type heterogeneity, and suggest these cell types provide a model system for exploring the organization and function of such heterogeneity. Given that the hippocampus is structurally simple and evolutionarily ancient, within-cell-type heterogeneity is likely to be a general and crucial feature of the mammalian brain.
A neuronal population encodes information most efficiently when its activity is uncorrelated and high-dimensional, and most robustly when its activity is correlated and lower-dimensional. Here, we analyzed the correlation structure of natural image coding, in large visual cortical populations recorded from awake mice. Evoked population activity was high dimensional, with correlations obeying an unexpected power-law: the n-th principal component variance scaled as 1/n. This was not inherited from the 1/f spectrum of natural images, because it persisted after stimulus whitening. We proved mathematically that the variance spectrum must decay at least this fast if a population code is smooth, i.e. if small changes in input cannot dominate population activity. The theory also predicts larger power-law exponents for lower-dimensional stimulus ensembles, which we validated experimentally. These results suggest that coding smoothness represents a fundamental constraint governing correlations in neural population codes.
Calcium imaging with genetically encoded calcium indicators (GECIs) is routinely used to measure neural activity in intact nervous systems. GECIs are frequently used in one of two different modes: to track activity in large populations of neuronal cell bodies, or to follow dynamics in subcellular compartments such as axons, dendrites and individual synaptic compartments. Despite major advances, calcium imaging is still limited by the biophysical properties of existing GECIs, including affinity, signal-to-noise ratio, rise and decay kinetics and dynamic range. Using structure-guided mutagenesis and neuron-based screening, we optimized the green fluorescent protein-based GECI GCaMP6 for different modes of in vivo imaging. The resulting jGCaMP7 sensors provide improved detection of individual spikes (jGCaMP7s,f), imaging in neurites and neuropil (jGCaMP7b), and may allow tracking larger populations of neurons using two-photon (jGCaMP7s,f) or wide-field (jGCaMP7c) imaging.
Noroviruses are a leading cause of foodborne illnesses worldwide. Although GII.4 strains have been responsible for most norovirus outbreaks, the assembled virus shell structures have been available in detail for only a single strain (GI.1). We present high-resolution (2.6- to 4.1-Å) cryoelectron microscopy (cryo-EM) structures of GII.4, GII.2, GI.7, and GI.1 human norovirus outbreak strain virus-like particles (VLPs). Although norovirus VLPs have been thought to exist in a single-sized assembly, our structures reveal polymorphism between and within genogroups, with small, medium, and large particle sizes observed. Using asymmetric reconstruction, we were able to resolve a Zn2+ metal ion adjacent to the coreceptor binding site, which affected the structural stability of the shell. Our structures serve as valuable templates for facilitating vaccine formulations.
In the spindle midzone, microtubules from opposite half-spindles form bundles between segregating chromosomes. Microtubule bundles can either push or restrict chromosome movement during anaphase in different cellular contexts, but how these activities are achieved remains poorly understood. Here, we use high-resolution live-cell imaging to analyze individual microtubule bundles, growing filaments, and chromosome movement in dividing human cells. Within bundles, filament overlap length marked by the cross-linking protein PRC1 decreases during anaphase as chromosome segregation slows. Filament ends within microtubule bundles appear capped despite dynamic PRC1 turnover and submicrometer proximity to growing microtubules. Chromosome segregation distance and rate are increased in two human cell lines when microtubule bundle assembly is prevented via PRC1 knockdown. Upon expressing a mutant PRC1 with reduced microtubule affinity, bundles assemble but chromosome hypersegregation is still observed. We propose that microtubule overlap length reduction, typically linked to pushing forces generated within filament bundles, is needed to properly restrict spindle elongation and position chromosomes within daughter cells.