Filter
Associated Lab
- Aso Lab (30) Apply Aso Lab filter
- Betzig Lab (1) Apply Betzig Lab filter
- Bock Lab (2) Apply Bock Lab filter
- Branson Lab (9) Apply Branson Lab filter
- Card Lab (5) Apply Card Lab filter
- Clapham Lab (1) Apply Clapham Lab filter
- Dickson Lab (2) Apply Dickson Lab filter
- Druckmann Lab (1) Apply Druckmann Lab filter
- Fetter Lab (1) Apply Fetter Lab filter
- Funke Lab (2) Apply Funke Lab filter
- Harris Lab (3) Apply Harris Lab filter
- Heberlein Lab (2) Apply Heberlein Lab filter
- Hermundstad Lab (2) Apply Hermundstad Lab filter
- Hess Lab (6) Apply Hess Lab filter
- Jayaraman Lab (7) Apply Jayaraman Lab filter
- Lippincott-Schwartz Lab (1) Apply Lippincott-Schwartz Lab filter
- Looger Lab (2) Apply Looger Lab filter
- O'Shea Lab (1) Apply O'Shea Lab filter
- Otopalik Lab (1) Apply Otopalik Lab filter
- Reiser Lab (17) Apply Reiser Lab filter
- Riddiford Lab (1) Apply Riddiford Lab filter
- Romani Lab (2) Apply Romani Lab filter
- Rubin Lab (148) Apply Rubin Lab filter
- Saalfeld Lab (5) Apply Saalfeld Lab filter
- Scheffer Lab (8) Apply Scheffer Lab filter
- Schreiter Lab (1) Apply Schreiter Lab filter
- Simpson Lab (3) Apply Simpson Lab filter
- Singer Lab (1) Apply Singer Lab filter
- Spruston Lab (1) Apply Spruston Lab filter
- Stern Lab (1) Apply Stern Lab filter
- Svoboda Lab (3) Apply Svoboda Lab filter
- Tjian Lab (1) Apply Tjian Lab filter
- Truman Lab (4) Apply Truman Lab filter
- Turaga Lab (1) Apply Turaga Lab filter
- Turner Lab (5) Apply Turner Lab filter
- Zuker Lab (1) Apply Zuker Lab filter
Associated Project Team
- CellMap (1) Apply CellMap filter
- Fly Functional Connectome (4) Apply Fly Functional Connectome filter
- Fly Olympiad (3) Apply Fly Olympiad filter
- FlyEM (12) Apply FlyEM filter
- FlyLight (20) Apply FlyLight filter
- GENIE (1) Apply GENIE filter
- Transcription Imaging (1) Apply Transcription Imaging filter
Publication Date
- 2025 (9) Apply 2025 filter
- 2024 (4) Apply 2024 filter
- 2023 (5) Apply 2023 filter
- 2022 (1) Apply 2022 filter
- 2021 (4) Apply 2021 filter
- 2020 (9) Apply 2020 filter
- 2019 (6) Apply 2019 filter
- 2018 (7) Apply 2018 filter
- 2017 (15) Apply 2017 filter
- 2016 (3) Apply 2016 filter
- 2015 (16) Apply 2015 filter
- 2014 (9) Apply 2014 filter
- 2013 (5) Apply 2013 filter
- 2012 (8) Apply 2012 filter
- 2011 (4) Apply 2011 filter
- 2010 (4) Apply 2010 filter
- 2009 (2) Apply 2009 filter
- 2008 (4) Apply 2008 filter
- 2007 (2) Apply 2007 filter
- 2006 (1) Apply 2006 filter
- 2002 (1) Apply 2002 filter
- 2000 (2) Apply 2000 filter
- 1999 (1) Apply 1999 filter
- 1997 (1) Apply 1997 filter
- 1995 (2) Apply 1995 filter
- 1994 (2) Apply 1994 filter
- 1993 (2) Apply 1993 filter
- 1992 (1) Apply 1992 filter
- 1991 (2) Apply 1991 filter
- 1990 (3) Apply 1990 filter
- 1989 (2) Apply 1989 filter
- 1987 (2) Apply 1987 filter
- 1986 (1) Apply 1986 filter
- 1985 (1) Apply 1985 filter
- 1984 (1) Apply 1984 filter
- 1983 (1) Apply 1983 filter
- 1982 (2) Apply 1982 filter
- 1981 (1) Apply 1981 filter
- 1979 (1) Apply 1979 filter
- 1973 (1) Apply 1973 filter
Type of Publication
148 Publications
Showing 41-50 of 148 results
A comparative analysis of the genomes of Drosophila melanogaster, Caenorhabditis elegans, and Saccharomyces cerevisiae-and the proteins they are predicted to encode-was undertaken in the context of cellular, developmental, and evolutionary processes. The nonredundant protein sets of flies and worms are similar in size and are only twice that of yeast, but different gene families are expanded in each genome, and the multidomain proteins and signaling pathways of the fly and worm are far more complex than those of yeast. The fly has orthologs to 177 of the 289 human disease genes examined and provides the foundation for rapid analysis of some of the basic processes involved in human disease.
Understanding the circuit mechanisms behind motion detection is a long-standing question in visual neuroscience. In , recent synapse-level connectomes in the optic lobe, particularly in ON-pathway (T4) receptive-field circuits, in concert with physiological studies, suggest an increasingly intricate motion model compared with the ubiquitous Hassenstein-Reichardt model, while our knowledge of OFF-pathway (T5) has been incomplete. Here we present a conclusive and comprehensive connectome that for the first time integrates detailed connectivity information for inputs to both T4 and T5 pathways in a single EM dataset covering the entire optic lobe. With novel reconstruction methods using automated synapse prediction suited to such a large connectome, we successfully corroborate previous findings in the T4 pathway and comprehensively identify inputs and receptive fields for T5. While the two pathways are likely evolutionarily linked and indeed exhibit many similarities, we uncover interesting differences and interactions that may underlie their distinct functional properties.
Nervous systems contain sensory neurons, local neurons, projection neurons, and motor neurons. To understand how these building blocks form whole circuits, we must distil these broad classes into neuronal cell types and describe their network connectivity. Using an electron micrograph dataset for an entire Drosophila melanogaster brain, we reconstruct the first complete inventory of olfactory projections connecting the antennal lobe, the insect analog of the mammalian olfactory bulb, to higher-order brain regions in an adult animal brain. We then connect this inventory to extant data in the literature, providing synaptic-resolution "holotypes" both for heavily investigated and previously unknown cell types. Projection neurons are approximately twice as numerous as reported by light level studies; cell types are stereotyped, but not identical, in cell and synapse numbers between brain hemispheres. The lateral horn, the insect analog of the mammalian cortical amygdala, is the main target for this olfactory information and has been shown to guide innate behavior. Here, we find new connectivity motifs, including axo-axonic connectivity between projection neurons, feedback, and lateral inhibition of these axons by a large population of neurons, and the convergence of different inputs, including non-olfactory inputs and memory-related feedback onto third-order olfactory neurons. These features are less prominent in the mushroom body calyx, the insect analog of the mammalian piriform cortex and a center for associative memory. Our work provides a complete neuroanatomical platform for future studies of the adult Drosophila olfactory system.
Vision provides animals with detailed information about their surroundings, conveying diverse features such as color, form, and movement across the visual scene. Computing these parallel spatial features requires a large and diverse network of neurons, such that in animals as distant as flies and humans, visual regions comprise half the brain’s volume. These visual brain regions often reveal remarkable structure-function relationships, with neurons organized along spatial maps with shapes that directly relate to their roles in visual processing. To unravel the stunning diversity of a complex visual system, a careful mapping of the neural architecture matched to tools for targeted exploration of that circuitry is essential. Here, we report a new connectome of the right optic lobe from a male Drosophila central nervous system FIB-SEM volume and a comprehensive inventory of the fly’s visual neurons. We developed a computational framework to quantify the anatomy of visual neurons, establishing a basis for interpreting how their shapes relate to spatial vision. By integrating this analysis with connectivity information, neurotransmitter identity, and expert curation, we classified the 53,000 neurons into 727 types, about half of which are systematically described and named for the first time. Finally, we share an extensive collection of split-GAL4 lines matched to our neuron type catalog. Together, this comprehensive set of tools and data unlock new possibilities for systematic investigations of vision in Drosophila, a foundation for a deeper understanding of sensory processing.
Many animals use visual information to navigate, but how such information is encoded and integrated by the navigation system remains incompletely understood. In Drosophila melanogaster, EPG neurons in the central complex compute the heading direction by integrating visual input from ER neurons, which are part of the anterior visual pathway (AVP). Here we densely reconstruct all neurons in the AVP using electron-microscopy data. The AVP comprises four neuropils, sequentially linked by three major classes of neurons: MeTu neurons, which connect the medulla in the optic lobe to the small unit of the anterior optic tubercle (AOTUsu) in the central brain; TuBu neurons, which connect the AOTUsu to the bulb neuropil; and ER neurons, which connect the bulb to the EPG neurons. On the basis of morphologies, connectivity between neural classes and the locations of synapses, we identify distinct information channels that originate from four types of MeTu neurons, and we further divide these into ten subtypes according to the presynaptic connections in the medulla and the postsynaptic connections in the AOTUsu. Using the connectivity of the entire AVP and the dendritic fields of the MeTu neurons in the optic lobes, we infer potential visual features and the visual area from which any ER neuron receives input. We confirm some of these predictions physiologically. These results provide a strong foundation for understanding how distinct sensory features can be extracted and transformed across multiple processing stages to construct higher-order cognitive representations.
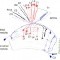
Motion detection is a fundamental neural computation performed by many sensory systems. In the fly, local motion computation is thought to occur within the first two layers of the visual system, the lamina and medulla. We constructed specific genetic driver lines for each of the 12 neuron classes in the lamina. We then depolarized and hyperpolarized each neuron type and quantified fly behavioral responses to a diverse set of motion stimuli. We found that only a small number of lamina output neurons are essential for motion detection, while most neurons serve to sculpt and enhance these feedforward pathways. Two classes of feedback neurons (C2 and C3), and lamina output neurons (L2 and L4), are required for normal detection of directional motion stimuli. Our results reveal a prominent role for feedback and lateral interactions in motion processing and demonstrate that motion-dependent behaviors rely on contributions from nearly all lamina neuron classes.
The Drosophila mushroom body (MB) is an associative learning network that is important for the control of sleep. We have recently identified particular intrinsic MB Kenyon cell (KC) classes that regulate sleep through synaptic activation of particular MB output neurons (MBONs) whose axons convey sleep control signals out of the MB to downstream target regions. Specifically, we found that sleep-promoting KCs increase sleep by preferentially activating cholinergic sleep-promoting MBONs, while wake-promoting KCs decrease sleep by preferentially activating glutamatergic wake-promoting MBONs. Here we use a combination of genetic and physiological approaches to identify wake-promoting dopaminergic neurons (DANs) that innervate the MB, and show that they activate wake-promoting MBONs. These studies reveal a dopaminergic sleep control mechanism that likely operates by modulation of KC-MBON microcircuits.
Optical and electron microscopy have made tremendous inroads toward understanding the complexity of the brain. However, optical microscopy offers insufficient resolution to reveal subcellular details, and electron microscopy lacks the throughput and molecular contrast to visualize specific molecular constituents over millimeter-scale or larger dimensions. We combined expansion microscopy and lattice light-sheet microscopy to image the nanoscale spatial relationships between proteins across the thickness of the mouse cortex or the entire Drosophila brain. These included synaptic proteins at dendritic spines, myelination along axons, and presynaptic densities at dopaminergic neurons in every fly brain region. The technology should enable statistically rich, large-scale studies of neural development, sexual dimorphism, degree of stereotypy, and structural correlations to behavior or neural activity, all with molecular contrast.
Previously, we identified that visual and olfactory associative memories of Drosophila share the mushroom body (MB) circuits (Vogt et al. 2014). Despite well-characterized odor representations in the Drosophila MB, the MB circuit for visual information is totally unknown. Here we show that a small subset of MB Kenyon cells (KCs) selectively responds to visual but not olfactory stimulation. The dendrites of these atypical KCs form a ventral accessory calyx (vAC), distinct from the main calyx that receives olfactory input. We identified two types of visual projection neurons (VPNs) directly connecting the optic lobes and the vAC. Strikingly, these VPNs are differentially required for visual memories of color and brightness. The segregation of visual and olfactory domains in the MB allows independent processing of distinct sensory memories and may be a conserved form of sensory representations among insects.

Visual motion perception is critical to many animal behaviors, and flies have emerged as a powerful model system for exploring this fundamental neural computation. Although numerous studies have suggested that fly motion vision is governed by a simple neural circuit [1-3], the implementation of this circuit has remained mysterious for decades. Connectomics and neurogenetics have produced a surge in recent progress, and several studies have shown selectivity for light increments (ON) or decrements (OFF) in key elements associated with this circuit [4-7]. However, related studies have reached disparate conclusions about where this selectivity emerges and whether it plays a major role in motion vision [8-13]. To address these questions, we examined activity in the neuropil thought to be responsible for visual motion detection, the medulla, of Drosophila melanogaster in response to a range of visual stimuli using two-photon calcium imaging. We confirmed that the input neurons of the medulla, the LMCs, are not responsible for light-on and light-off selectivity. We then examined the pan-neural response of medulla neurons and found prominent selectivity for light-on and light-off in layers of the medulla associated with two anatomically derived pathways (L1/L2 associated) [14, 15]. We next examined the activity of prominent interneurons within each pathway (Mi1 and Tm1) and found that these neurons have corresponding selectivity for light-on or light-off. These results provide direct evidence that motion is computed in parallel light-on and light-off pathways, demonstrate that this selectivity emerges in neurons immediately downstream of the LMCs, and specify where crucial elements of motion computation occur.