Filter
Associated Lab
- Betzig Lab (2) Apply Betzig Lab filter
- Chklovskii Lab (4) Apply Chklovskii Lab filter
- Eddy/Rivas Lab (6) Apply Eddy/Rivas Lab filter
- Fetter Lab (2) Apply Fetter Lab filter
- Hess Lab (2) Apply Hess Lab filter
- Jayaraman Lab (1) Apply Jayaraman Lab filter
- Leonardo Lab (1) Apply Leonardo Lab filter
- Looger Lab (7) Apply Looger Lab filter
- Magee Lab (3) Apply Magee Lab filter
- Menon Lab (1) Apply Menon Lab filter
- Reiser Lab (1) Apply Reiser Lab filter
- Riddiford Lab (4) Apply Riddiford Lab filter
- Rubin Lab (1) Apply Rubin Lab filter
- Schreiter Lab (2) Apply Schreiter Lab filter
- Shroff Lab (2) Apply Shroff Lab filter
- Simpson Lab (1) Apply Simpson Lab filter
- Spruston Lab (1) Apply Spruston Lab filter
- Sternson Lab (1) Apply Sternson Lab filter
- Svoboda Lab (8) Apply Svoboda Lab filter
- Truman Lab (2) Apply Truman Lab filter
Associated Support Team
Publication Date
- December 2009 (5) Apply December 2009 filter
- November 2009 (2) Apply November 2009 filter
- October 2009 (7) Apply October 2009 filter
- September 2009 (3) Apply September 2009 filter
- August 2009 (2) Apply August 2009 filter
- July 2009 (7) Apply July 2009 filter
- June 2009 (4) Apply June 2009 filter
- May 2009 (4) Apply May 2009 filter
- April 2009 (2) Apply April 2009 filter
- March 2009 (5) Apply March 2009 filter
- February 2009 (4) Apply February 2009 filter
- January 2009 (11) Apply January 2009 filter
- Remove 2009 filter 2009
56 Janelia Publications
Showing 21-30 of 56 resultsCortical information processing is under state-dependent control of subcortical neuromodulatory systems. Although this modulatory effect is thought to be mediated mainly by slow nonsynaptic metabotropic receptors, other mechanisms, such as direct synaptic transmission, are possible. Yet, it is currently unknown if any such form of subcortical control exists. Here, we present direct evidence of a strong, spatiotemporally precise excitatory input from an ascending neuromodulatory center. Selective stimulation of serotonergic median raphe neurons produced a rapid activation of hippocampal interneurons. At the network level, this subcortical drive was manifested as a pattern of effective disynaptic GABAergic inhibition that spread throughout the circuit. This form of subcortical network regulation should be incorporated into current concepts of normal and pathological cortical function.
Imaging informatics has emerged as a major research theme in biomedicine in the last few decades. Currently, personalised, predictive and preventive patient care is believed to be one of the top priorities in biomedical research and practice. Imaging informatics plays a major role in biomedicine studies. This paper reviews main applications and challenges of imaging informatics in biomedicine.
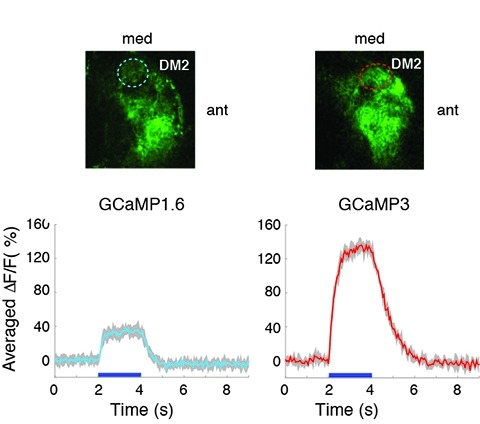
Genetically encoded calcium indicators (GECIs) can be used to image activity in defined neuronal populations. However, current GECIs produce inferior signals compared to synthetic indicators and recording electrodes, precluding detection of low firing rates. We developed a single-wavelength GCaMP2-based GECI (GCaMP3), with increased baseline fluorescence (3-fold), increased dynamic range (3-fold) and higher affinity for calcium (1.3-fold). We detected GCaMP3 fluorescence changes triggered by single action potentials in pyramidal cell dendrites, with signal-to-noise ratio and photostability substantially better than those of GCaMP2, D3cpVenus and TN-XXL. In Caenorhabditis elegans chemosensory neurons and the Drosophila melanogaster antennal lobe, sensory stimulation-evoked fluorescence responses were significantly enhanced with GCaMP3 (4-6-fold). In somatosensory and motor cortical neurons in the intact mouse, GCaMP3 detected calcium transients with amplitudes linearly dependent on action potential number. Long-term imaging in the motor cortex of behaving mice revealed large fluorescence changes in imaged neurons over months.
SUMMARY: INFERNAL builds consensus RNA secondary structure profiles called covariance models (CMs), and uses them to search nucleic acid sequence databases for homologous RNAs, or to create new sequence- and structure-based multiple sequence alignments. AVAILABILITY: Source code, documentation and benchmark downloadable from http://infernal.janelia.org. INFERNAL is freely licensed under the GNU GPLv3 and should be portable to any POSIX-compliant operating system, including Linux and Mac OS/X.
Understanding molecular-scale architecture of cells requires determination of 3D locations of specific proteins with accuracy matching their nanometer-length scale. Existing electron and light microscopy techniques are limited either in molecular specificity or resolution. Here, we introduce interferometric photoactivated localization microscopy (iPALM), the combination of photoactivated localization microscopy with single-photon, simultaneous multiphase interferometry that provides sub-20-nm 3D protein localization with optimal molecular specificity. We demonstrate measurement of the 25-nm microtubule diameter, resolve the dorsal and ventral plasma membranes, and visualize the arrangement of integrin receptors within endoplasmic reticulum and adhesion complexes, 3D protein organization previously resolved only by electron microscopy. iPALM thus closes the gap between electron tomography and light microscopy, enabling both molecular specification and resolution of cellular nanoarchitecture.
Although the vinegar fly, Drosophila melanogaster, has been a biological model organism for over a century, its emergence as a model system for the study of neurophysiology is comparatively recent. The primary reason for this is that the vinegar fly and its neurons are tiny; up until 5 years ago, it was prohibitively difficult to record intracellularly from individual neurons in the intact Drosophila brain (Wilson et al., 2004). Today, fly electrophysiologists can genetically label neurons with GFP and reliably record from many (but not all) neurons in the fruit fly brain. Using genetic tools to drive expression of fluorescent calcium indicators, light-sensitive ion channels, or cell activity suppressors, we are beginning to understand how the external environment is represented with electrical potentials in Drosophila neurons (for review, see Olsen and Wilson, 2008).
Accuracy of automated structural RNA alignment is improved by using models that consider not only primary sequence but also secondary structure information. However, current RNA structural alignment approaches tend to perform poorly on incomplete sequence fragments, such as single reads from metagenomic environmental surveys, because nucleotides that are expected to be base paired are missing.
A low-contrast spot that activates just one ganglion cell in the retina is detected in the spike train of the cell with about the same sensitivity as it is detected behaviorally. This is consistent with Barlow’s proposal that the ganglion cell and later stages of spiking neurons transfer information essentially without loss. Yet, when losses of sensitivity by all preneural factors are accounted for, predicted sensitivity near threshold is considerably greater than behavioral sensitivity, implying that somewhere in the brain information is lost. We hypothesized that the losses occur mainly in the retina, where graded signals are processed by analog circuits that transfer information at high rates and low metabolic cost. To test this, we constructed a model that included all preneural losses for an in vitro mammalian retina, and evaluated the model to predict sensitivity at the cone output. Recording graded responses postsynaptic to the cones (from the type A horizontal cell) and comparing to predicted preneural sensitivity, we found substantial loss of sensitivity (4.2-fold) across the first visual synapse. Recording spike responses from brisk-transient ganglion cells stimulated with the same spot, we found a similar loss (3.5-fold) across the second synapse. The total retinal loss approximated the known overall loss, supporting the hypothesis that from stimulus to perception, most loss near threshold is retinal.
Drosophila is a marvelous system to study the underlying principles that govern how neural circuits govern behaviors. The scale of the fly brain (approximately 100,000 neurons) and the complexity of the behaviors the fly can perform make it a tractable experimental model organism. In addition, 100 years and hundreds of labs have contributed to an extensive array of tools and techniques that can be used to dissect the function and organization of the fly nervous system. This review discusses both the conceptual challenges and the specific tools for a neurogenetic approach to circuit mapping in Drosophila.