Filter
Associated Lab
- Ahrens Lab (3) Apply Ahrens Lab filter
- Aso Lab (6) Apply Aso Lab filter
- Baker Lab (2) Apply Baker Lab filter
- Betzig Lab (11) Apply Betzig Lab filter
- Branson Lab (6) Apply Branson Lab filter
- Cardona Lab (5) Apply Cardona Lab filter
- Chklovskii Lab (2) Apply Chklovskii Lab filter
- Cui Lab (5) Apply Cui Lab filter
- Dickson Lab (1) Apply Dickson Lab filter
- Druckmann Lab (3) Apply Druckmann Lab filter
- Dudman Lab (3) Apply Dudman Lab filter
- Eddy/Rivas Lab (4) Apply Eddy/Rivas Lab filter
- Egnor Lab (1) Apply Egnor Lab filter
- Fetter Lab (5) Apply Fetter Lab filter
- Freeman Lab (7) Apply Freeman Lab filter
- Funke Lab (1) Apply Funke Lab filter
- Gonen Lab (5) Apply Gonen Lab filter
- Grigorieff Lab (7) Apply Grigorieff Lab filter
- Harris Lab (7) Apply Harris Lab filter
- Heberlein Lab (1) Apply Heberlein Lab filter
- Hess Lab (7) Apply Hess Lab filter
- Jayaraman Lab (4) Apply Jayaraman Lab filter
- Ji Lab (4) Apply Ji Lab filter
- Karpova Lab (1) Apply Karpova Lab filter
- Keleman Lab (2) Apply Keleman Lab filter
- Keller Lab (7) Apply Keller Lab filter
- Lavis Lab (5) Apply Lavis Lab filter
- Leonardo Lab (2) Apply Leonardo Lab filter
- Liu (Zhe) Lab (4) Apply Liu (Zhe) Lab filter
- Looger Lab (9) Apply Looger Lab filter
- Magee Lab (5) Apply Magee Lab filter
- Murphy Lab (1) Apply Murphy Lab filter
- Pastalkova Lab (3) Apply Pastalkova Lab filter
- Reiser Lab (2) Apply Reiser Lab filter
- Romani Lab (2) Apply Romani Lab filter
- Rubin Lab (16) Apply Rubin Lab filter
- Saalfeld Lab (3) Apply Saalfeld Lab filter
- Scheffer Lab (2) Apply Scheffer Lab filter
- Schreiter Lab (4) Apply Schreiter Lab filter
- Shroff Lab (1) Apply Shroff Lab filter
- Simpson Lab (4) Apply Simpson Lab filter
- Singer Lab (10) Apply Singer Lab filter
- Spruston Lab (7) Apply Spruston Lab filter
- Stern Lab (4) Apply Stern Lab filter
- Sternson Lab (7) Apply Sternson Lab filter
- Svoboda Lab (9) Apply Svoboda Lab filter
- Tjian Lab (6) Apply Tjian Lab filter
- Truman Lab (6) Apply Truman Lab filter
- Turaga Lab (1) Apply Turaga Lab filter
- Turner Lab (2) Apply Turner Lab filter
- Wu Lab (1) Apply Wu Lab filter
- Zlatic Lab (4) Apply Zlatic Lab filter
- Zuker Lab (2) Apply Zuker Lab filter
Associated Project Team
Associated Support Team
- Electron Microscopy (1) Apply Electron Microscopy filter
- Fly Facility (1) Apply Fly Facility filter
- Gene Targeting and Transgenics (3) Apply Gene Targeting and Transgenics filter
- Janelia Experimental Technology (2) Apply Janelia Experimental Technology filter
- Management Team (1) Apply Management Team filter
- Molecular Genomics (1) Apply Molecular Genomics filter
- Primary & iPS Cell Culture (2) Apply Primary & iPS Cell Culture filter
- Project Technical Resources (1) Apply Project Technical Resources filter
- Scientific Computing Software (11) Apply Scientific Computing Software filter
Publication Date
- December 2015 (15) Apply December 2015 filter
- November 2015 (22) Apply November 2015 filter
- October 2015 (16) Apply October 2015 filter
- September 2015 (16) Apply September 2015 filter
- August 2015 (17) Apply August 2015 filter
- July 2015 (18) Apply July 2015 filter
- June 2015 (16) Apply June 2015 filter
- May 2015 (16) Apply May 2015 filter
- April 2015 (18) Apply April 2015 filter
- March 2015 (16) Apply March 2015 filter
- February 2015 (15) Apply February 2015 filter
- January 2015 (10) Apply January 2015 filter
- Remove 2015 filter 2015
195 Janelia Publications
Showing 41-50 of 195 results
Imaging fast cellular dynamics across large specimens requires high resolution in all dimensions, high imaging speeds, good physical coverage and low photo-damage. To meet these requirements, we developed isotropic multiview (IsoView) light-sheet microscopy, which rapidly images large specimens via simultaneous light-sheet illumination and fluorescence detection along four orthogonal directions. Combining these four views by means of high-throughput multiview deconvolution yields images with high resolution in all three dimensions. We demonstrate whole-animal functional imaging of Drosophila larvae at a spatial resolution of 1.1-2.5 μm and temporal resolution of 2 Hz for several hours. We also present spatially isotropic whole-brain functional imaging in Danio rerio larvae and spatially isotropic multicolor imaging of fast cellular dynamics across gastrulating Drosophila embryos. Compared with conventional light-sheet microscopy, IsoView microscopy improves spatial resolution at least sevenfold and decreases resolution anisotropy at least threefold. Compared with existing high-resolution light-sheet techniques, IsoView microscopy effectively doubles the penetration depth and provides subsecond temporal resolution for specimens 400-fold larger than could previously be imaged.
To investigate the fundamental question of how nervous systems encode, organize, and sequence behaviors, Kato et al. imaged neural activity with cellular resolution across the brain of the worm Caenorhabditis elegans. Locomotion behavior seems to be continuously represented by cyclical patterns of distributed neural activity that are present even in immobilized animals.
Bilaterally symmetric motor patterns-those in which left-right pairs of muscles contract synchronously and with equal amplitude (such as breathing, smiling, whisking, and locomotion)-are widespread throughout the animal kingdom. Yet, surprisingly little is known about the underlying neural circuits. We performed a thermogenetic screen to identify neurons required for bilaterally symmetric locomotion in Drosophila larvae and identified the evolutionarily conserved Even-skipped(+) interneurons (Eve/Evx). Activation or ablation of Eve(+) interneurons disrupted bilaterally symmetric muscle contraction amplitude, without affecting the timing of motor output. Eve(+) interneurons are not rhythmically active and thus function independently of the locomotor CPG. GCaMP6 calcium imaging of Eve(+) interneurons in freely moving larvae showed left-right asymmetric activation that correlated with larval behavior. TEM reconstruction of Eve(+) interneuron inputs and outputs showed that the Eve(+) interneurons are at the core of a sensorimotor circuit capable of detecting and modifying body wall muscle contraction.
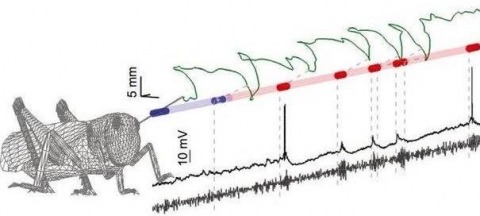
Sensory inputs are often fluctuating and intermittent, yet animals reliably utilize them to direct behavior. Here we ask how natural stimulus fluctuations influence the dynamic neural encoding of odors. Using the locust olfactory system, we isolated two main causes of odor intermittency: chaotic odor plumes and active sampling behaviors. Despite their irregularity, chaotic odor plumes still drove dynamic neural response features including the synchronization, temporal patterning, and short-term plasticity of spiking in projection neurons, enabling classifier-based stimulus identification and activating downstream decoders (Kenyon cells). Locusts can also impose odor intermittency through active sampling movements with their unrestrained antennae. Odors triggered immediate, spatially targeted antennal scanning that, paradoxically, weakened individual neural responses. However, these frequent but weaker responses were highly informative about stimulus location. Thus, not only are odor-elicited dynamic neural responses compatible with natural stimulus fluctuations and important for stimulus identification, but locusts actively increase intermittency, possibly to improve stimulus localization.
Early transplantation and grafting experiments suggest that body organs follow autonomous growth programs [1-3], therefore pointing to a need for coordination mechanisms to produce fit individuals with proper proportions. We recently identified Drosophila insulin-like peptide 8 (Dilp8) as a relaxin and insulin-like molecule secreted from growing tissues that plays a central role in coordinating growth between organs and coupling organ growth with animal maturation [4, 5]. Deciphering the function of Dilp8 in growth coordination relies on the identification of the receptor and tissues relaying Dilp8 signaling. We show here that the orphan receptor leucine-rich repeat-containing G protein-coupled receptor 3 (Lgr3), a member of the highly conserved family of relaxin family peptide receptors (RXFPs), mediates the checkpoint function of Dilp8 for entry into maturation. We functionally identify two Lgr3-positive neurons in each brain lobe that are required to induce a developmental delay upon overexpression of Dilp8. These neurons are located in the pars intercerebralis, an important neuroendocrine area in the brain, and make physical contacts with the PTTH neurons that ultimately control the production and release of the molting steroid ecdysone. Reducing Lgr3 levels in these neurons results in adult flies exhibiting increased fluctuating bilateral asymmetry, therefore recapitulating the phenotype of dilp8 mutants. Our work reveals a novel Dilp8/Lgr3 neuronal circuitry involved in a feedback mechanism that ensures coordination between organ growth and developmental transitions and prevents developmental variability.
The frequencies of transcription initiation of regulated and constitutive genes depend on the concentration of free RNA polymerase holoenzyme [Rf] near their promoters. Although RNA polymerase is largely confined to the nucleoid, it is difficult to determine absolute concentrations of [Rf] at particular locations within the nucleoid structure. However, relative concentrations of free RNA polymerase at different growth rates, [Rf]rel, can be estimated from the activities of constitutive promoters. Previous studies indicated that the rrnB P2 promoter is constitutive and that [Rf]rel in the vicinity of rrnB P2 increases with increasing growth rate. Recently it has become possible to directly visualize Rf in growing Escherichia coli cells. Here we examine some of the important issues relating to gene expression based on these new observations. We conclude that: (i) At a growth rate of 2 doublings/h, there are about 1000 free and 2350 non-specifically DNA-bound RNA polymerase molecules per average cell (12 and 28%, respectively, of 8400 total) which are in rapid equilibrium. (ii) The reversibility of the non-specific binding generates more than 1000 free RNA polymerase molecules every second in the immediate vicinity of the DNA. Of these, most rebind non-specifically to the DNA within a few ms; the frequency of non-specific binding is at least two orders of magnitude greater than specific binding and transcript initiation. (iii) At a given amount of RNA polymerase per cell, [Rf] and the density of non-specifically DNA-bound RNA polymerase molecules along the DNA both vary reciprocally with the amount of DNA in the cell. (iv) At 2 doublings/h an E. coli cell contains, on the average, about 1 non-specifically bound RNA polymerase per 9 kbp of DNA and 1 free RNA polymerase per 20 kbp of DNA. However some DNA regions (i.e. near active rRNA operons) may have significantly higher than average [Rf].
Neural stem cells show age-dependent developmental potentials, as evidenced by their production of distinct neuron types at different developmental times. Drosophila neuroblasts produce long, stereotyped lineages of neurons. We searched for factors that could regulate neural temporal fate by RNA-sequencing lineage-specific neuroblasts at various developmental times. We found that two RNA-binding proteins, IGF-II mRNA-binding protein (Imp) and Syncrip (Syp), display opposing high-to-low and low-to-high temporal gradients with lineage-specific temporal dynamics. Imp and Syp promote early and late fates, respectively, in both a slowly progressing and a rapidly changing lineage. Imp and Syp control neuronal fates in the mushroom body lineages by regulating the temporal transcription factor Chinmo translation. Together, the opposing Imp/Syp gradients encode stem cell age, specifying multiple cell fates within a lineage.
Far-field optical microscopy using focused light is an important tool in a number of scientific disciplines including chemical, (bio)physical and biomedical research, particularly with respect to the study of living cells and organisms. Unfortunately, the applicability of the optical microscope is limited, since the diffraction of light imposes limitations on the spatial resolution of the image. Consequently the details of, for example, cellular protein distributions, can be visualized only to a certain extent. Fortunately, recent years have witnessed the development of 'super-resolution' far-field optical microscopy (nanoscopy) techniques such as stimulated emission depletion (STED), ground state depletion (GSD), reversible saturated optical (fluorescence) transitions (RESOLFT), photoactivation localization microscopy (PALM), stochastic optical reconstruction microscopy (STORM), structured illumination microscopy (SIM) or saturated structured illumination microscopy (SSIM), all in one way or another addressing the problem of the limited spatial resolution of far-field optical microscopy. While SIM achieves a two-fold improvement in spatial resolution compared to conventional optical microscopy, STED, RESOLFT, PALM/STORM, or SSIM have all gone beyond, pushing the limits of optical image resolution to the nanometer scale. Consequently, all super-resolution techniques open new avenues of biomedical research. Because the field is so young, the potential capabilities of different super-resolution microscopy approaches have yet to be fully explored, and uncertainties remain when considering the best choice of methodology. Thus, even for experts, the road to the future is sometimes shrouded in mist. The super-resolution optical microscopy roadmap of Journal of Physics D: Applied Physicsaddresses this need for clarity. It provides guidance to the outstanding questions through a collection of short review articles from experts in the field, giving a thorough discussion on the concepts underlying super-resolution optical microscopy, the potential of different approaches, the importance of label optimization (such as reversible photoswitchable proteins) and applications in which these methods will have a significant impact.
Endoplasmic reticulum calcium homeostasis is critical for cellular functions and is disrupted in diverse pathologies including neurodegeneration and cardiovascular disease. Owing to the high concentration of calcium within the ER, studying this subcellular compartment requires tools that are optimized for these conditions. To develop a single-fluorophore genetically encoded calcium indicator for this organelle, we targeted a low affinity variant of GCaMP3 to the ER lumen (GCaMPer (10.19)). A set of viral vectors was constructed to express GCaMPer in human neuroblastoma cells, rat primary cortical neurons, and human induced pluripotent stem cell-derived cardiomyocytes. We observed dynamic changes in GCaMPer (10.19) fluorescence in response to pharmacologic manipulations of the ER calcium store. Additionally, periodic calcium efflux from the ER was observed during spontaneous beating of cardiomyocytes. GCaMPer (10.19) has utility in imaging ER calcium in living cells and providing insight into luminal calcium dynamics under physiologic and pathologic states.
Animal sensory systems are optimally adapted to those features typically encountered in natural surrounds, thus allowing neurons with limited bandwidth to encode challengingly large input ranges. Natural scenes are not random, and peripheral visual systems in vertebrates and insects have evolved to respond efficiently to their typical spatial statistics. The mammalian visual cortex is also tuned to natural spatial statistics, but less is known about coding in higher order neurons in insects. To redress this we here record intracellularly from a higher order visual neuron in the hoverfly. We show that the cSIFE neuron, which is inhibited by stationary images, is maximally inhibited when the slope constant of the amplitude spectrum is close to the mean in natural scenes. The behavioural optomotor response is also strongest to images with naturalistic image statistics. Our results thus reveal a close coupling between the inherent statistics of natural scenes and higher order visual processing in insects.