Filter
Associated Lab
- Aso Lab (1) Apply Aso Lab filter
- Baker Lab (3) Apply Baker Lab filter
- Betzig Lab (7) Apply Betzig Lab filter
- Bock Lab (1) Apply Bock Lab filter
- Cui Lab (3) Apply Cui Lab filter
- Dickson Lab (1) Apply Dickson Lab filter
- Dudman Lab (1) Apply Dudman Lab filter
- Eddy/Rivas Lab (5) Apply Eddy/Rivas Lab filter
- Fetter Lab (2) Apply Fetter Lab filter
- Gonen Lab (1) Apply Gonen Lab filter
- Hess Lab (2) Apply Hess Lab filter
- Jayaraman Lab (2) Apply Jayaraman Lab filter
- Ji Lab (2) Apply Ji Lab filter
- Keller Lab (2) Apply Keller Lab filter
- Lavis Lab (5) Apply Lavis Lab filter
- Lee (Albert) Lab (1) Apply Lee (Albert) Lab filter
- Leonardo Lab (2) Apply Leonardo Lab filter
- Looger Lab (7) Apply Looger Lab filter
- Magee Lab (1) Apply Magee Lab filter
- Menon Lab (3) Apply Menon Lab filter
- Murphy Lab (1) Apply Murphy Lab filter
- Reiser Lab (2) Apply Reiser Lab filter
- Riddiford Lab (1) Apply Riddiford Lab filter
- Rubin Lab (3) Apply Rubin Lab filter
- Scheffer Lab (2) Apply Scheffer Lab filter
- Schreiter Lab (2) Apply Schreiter Lab filter
- Simpson Lab (3) Apply Simpson Lab filter
- Singer Lab (1) Apply Singer Lab filter
- Sternson Lab (6) Apply Sternson Lab filter
- Svoboda Lab (7) Apply Svoboda Lab filter
- Tjian Lab (2) Apply Tjian Lab filter
- Truman Lab (1) Apply Truman Lab filter
- Zlatic Lab (1) Apply Zlatic Lab filter
- Zuker Lab (2) Apply Zuker Lab filter
Associated Project Team
Publication Date
- December 2011 (10) Apply December 2011 filter
- November 2011 (8) Apply November 2011 filter
- October 2011 (8) Apply October 2011 filter
- September 2011 (8) Apply September 2011 filter
- August 2011 (9) Apply August 2011 filter
- July 2011 (5) Apply July 2011 filter
- June 2011 (10) Apply June 2011 filter
- May 2011 (6) Apply May 2011 filter
- April 2011 (5) Apply April 2011 filter
- March 2011 (6) Apply March 2011 filter
- February 2011 (8) Apply February 2011 filter
- January 2011 (15) Apply January 2011 filter
- Remove 2011 filter 2011
98 Janelia Publications
Showing 21-30 of 98 resultsOlfactory systems encode odours by which neurons respond and by when they respond. In mammals, every sniff evokes a precise, odour-specific sequence of activity across olfactory neurons. Likewise, in a variety of neural systems, ranging from sensory periphery to cognitive centres, neuronal activity is timed relative to sampling behaviour and/or internally generated oscillations. As in these neural systems, relative timing of activity may represent information in the olfactory system. However, there is no evidence that mammalian olfactory systems read such cues. To test whether mice perceive the timing of olfactory activation relative to the sniff cycle (’sniff phase’), we used optogenetics in gene-targeted mice to generate spatially constant, temporally controllable olfactory input. Here we show that mice can behaviourally report the sniff phase of optogenetically driven activation of olfactory sensory neurons. Furthermore, mice can discriminate between light-evoked inputs that are shifted in the sniff cycle by as little as 10 milliseconds, which is similar to the temporal precision of olfactory bulb odour responses. Electrophysiological recordings in the olfactory bulb of awake mice show that individual cells encode the timing of photoactivation in relation to the sniff in both the timing and the amplitude of their responses. Our work provides evidence that the mammalian olfactory system can read temporal patterns, and suggests that timing of activity relative to sampling behaviour is a potent cue that may enable accurate olfactory percepts to form quickly.
In the rodent vibrissal system, active sensation and sensorimotor integration are mediated in part by connections between barrel cortex and vibrissal motor cortex. Little is known about how these structures interact at the level of neurons. We used Channelrhodopsin-2 (ChR2) expression, combined with anterograde and retrograde labeling, to map connections between barrel cortex and pyramidal neurons in mouse motor cortex. Barrel cortex axons preferentially targeted upper layer (L2/3, L5A) neurons in motor cortex; input to neurons projecting back to barrel cortex was particularly strong. Barrel cortex input to deeper layers (L5B, L6) of motor cortex, including neurons projecting to the brainstem, was weak, despite pronounced geometric overlap of dendrites with axons from barrel cortex. Neurons in different layers received barrel cortex input within stereotyped dendritic domains. The cortico-cortical neurons in superficial layers of motor cortex thus couple motor and sensory signals and might mediate sensorimotor integration and motor learning.
Mitral/tufted cells of the olfactory bulb receive odorant information from receptor neurons and transmit this information to the cortex. Studies in awake behaving animals have found that sustained responses of mitral cells to odorants are rare, suggesting sparse combinatorial representation of the odorants. Careful alignment of mitral cell firing with the phase of the respiration cycle revealed brief transient activity in the larger population of mitral cells, which respond to odorants during a small fraction of the respiration cycle. Responses of these cells are therefore temporally sparse. Here, we propose a mathematical model for the olfactory bulb network that can reproduce both combinatorially and temporally sparse mitral cell codes. We argue that sparse codes emerge as a result of the balance between mitral cells’ excitatory inputs and inhibition provided by the granule cells. Our model suggests functional significance for the dendrodendritic synapses mediating interactions between mitral and granule cells.
Profile hidden Markov models (profile HMMs) and probabilistic inference methods have made important contributions to the theory of sequence database homology search. However, practical use of profile HMM methods has been hindered by the computational expense of existing software implementations. Here I describe an acceleration heuristic for profile HMMs, the "multiple segment Viterbi" (MSV) algorithm. The MSV algorithm computes an optimal sum of multiple ungapped local alignment segments using a striped vector-parallel approach previously described for fast Smith/Waterman alignment. MSV scores follow the same statistical distribution as gapped optimal local alignment scores, allowing rapid evaluation of significance of an MSV score and thus facilitating its use as a heuristic filter. I also describe a 20-fold acceleration of the standard profile HMM Forward/Backward algorithms using a method I call "sparse rescaling". These methods are assembled in a pipeline in which high-scoring MSV hits are passed on for reanalysis with the full HMM Forward/Backward algorithm. This accelerated pipeline is implemented in the freely available HMMER3 software package. Performance benchmarks show that the use of the heuristic MSV filter sacrifices negligible sensitivity compared to unaccelerated profile HMM searches. HMMER3 is substantially more sensitive and 100- to 1000-fold faster than HMMER2. HMMER3 is now about as fast as BLAST for protein searches.
Full reconstruction of neuron morphology is of fundamental interest for the analysis and understanding of their functioning. We have developed a novel method capable of automatically tracing neurons in three-dimensional microscopy data. In contrast to template-based methods, the proposed approach makes no assumptions about the shape or appearance of neurite structure. Instead, an efficient seeding approach is applied to capture complex neuronal structures and the tracing problem is solved by computing the optimal reconstruction with a weighted graph. The optimality is determined by the cost function designed for the path between each pair of seeds and by topological constraints defining the component interrelations and completeness. In addition, an automated neuron comparison method is introduced for performance evaluation and structure analysis. The proposed algorithm is computationally efficient and has been validated using different types of microscopy data sets including Drosophila’s projection neurons and fly neurons with presynaptic sites. In all cases, the approach yielded promising results.
Research on the biology of addiction has advanced significantly over the last 50 years expanding our understanding of the brain mechanisms underlying reward, reinforcement and craving. Novel experimental approaches and techniques have provided an ever increasing armory of tools to dissect behavioral processes, neural networks and molecular mechanisms. The ultimate goal is to reintegrate this knowledge into a coherent, mechanistic framework of addiction to help identify new treatment. This can be greatly facilitated by using tools that allow, with great spatial and temporal specificity, to link molecular changes with altered activation of neural circuits and behavior. Such specificity can now be achieved by using optogenetic tools. Our review describes the general principles of optogenetics and its use to understand the links between neural activity and behavior. We also provide an overview of recent studies using optogenetic tools in addiction and consider some outstanding questions of addiction research that are particularly amenable for optogenetic approaches.
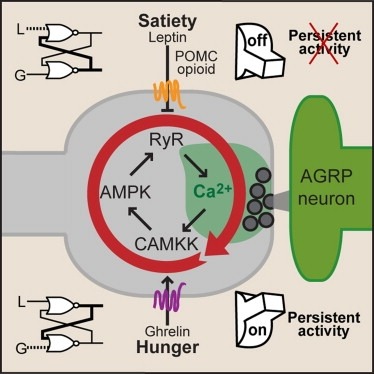
Synaptic plasticity in response to changes in physiologic state is coordinated by hormonal signals across multiple neuronal cell types. Here, we combine cell-type-specific electrophysiological, pharmacological, and optogenetic techniques to dissect neural circuits and molecular pathways controlling synaptic plasticity onto AGRP neurons, a population that regulates feeding. We find that food deprivation elevates excitatory synaptic input, which is mediated by a presynaptic positive feedback loop involving AMP-activated protein kinase. Potentiation of glutamate release was triggered by the orexigenic hormone ghrelin and exhibited hysteresis, persisting for hours after ghrelin removal. Persistent activity was reversed by the anorexigenic hormone leptin, and optogenetic photostimulation demonstrated involvement of opioid release from POMC neurons. Based on these experiments, we propose a memory storage device for physiological state constructed from bistable synapses that are flipped between two sustained activity states by transient exposure to hormones signaling energy levels.
A specialist neuron uses an intriguing process to help control the body's response to hunger. A lipid pathway involving the breakdown of cellular components regulates the expression of a neuropeptide that affects feeding and body weight.
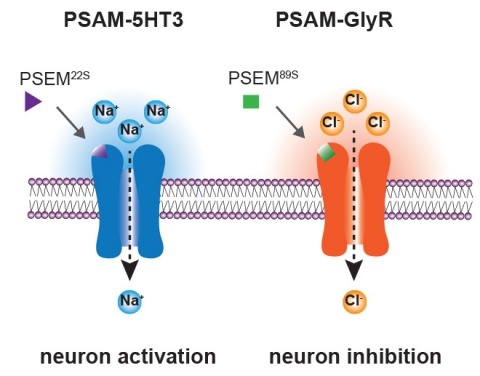
Ionic flux mediates essential physiological and behavioral functions in defined cell populations. Cell type-specific activators of diverse ionic conductances are needed for probing these effects. We combined chemistry and protein engineering to enable the systematic creation of a toolbox of ligand-gated ion channels (LGICs) with orthogonal pharmacologic selectivity and divergent functional properties. The LGICs and their small-molecule effectors were able to activate a range of ionic conductances in genetically specified cell types. LGICs constructed for neuronal perturbation could be used to selectively manipulate neuron activity in mammalian brains in vivo. The diversity of ion channel tools accessible from this approach will be useful for examining the relationship between neuronal activity and animal behavior, as well as for cell biological and physiological applications requiring chemical control of ion conductance.
Concomitant with the publication of this Special Issue of Neuroinformatics, a substantially updated version of the DIADEM web site has been released at http://diademchallenge.org. This web site was originally designed to host the challenge for automating the digital reconstruction of axonal and dendritic morphology (hence the DIADEM acronym). This post-competition version features additional content for continued use as the access point for DIADEM-related material. From the very beginning, one of the spirits of DIADEM has been to share data and resources with the neuroscience research community at large. The resources available from or linked to the DIADEM website constitute a substantial scientific legacy of the 2009/2010 competition. The new content includes finalist algorithms, image stack data, gold standard reconstructions, an updated DIADEM metric, and a retrospective on the competition in text and images.