Filter
Associated Lab
- Ahrens Lab (1) Apply Ahrens Lab filter
- Aso Lab (2) Apply Aso Lab filter
- Baker Lab (2) Apply Baker Lab filter
- Betzig Lab (5) Apply Betzig Lab filter
- Bock Lab (1) Apply Bock Lab filter
- Branson Lab (3) Apply Branson Lab filter
- Card Lab (1) Apply Card Lab filter
- Cardona Lab (6) Apply Cardona Lab filter
- Chklovskii Lab (1) Apply Chklovskii Lab filter
- Cui Lab (6) Apply Cui Lab filter
- Dickson Lab (3) Apply Dickson Lab filter
- Druckmann Lab (3) Apply Druckmann Lab filter
- Eddy/Rivas Lab (1) Apply Eddy/Rivas Lab filter
- Fetter Lab (1) Apply Fetter Lab filter
- Fitzgerald Lab (1) Apply Fitzgerald Lab filter
- Gonen Lab (7) Apply Gonen Lab filter
- Grigorieff Lab (7) Apply Grigorieff Lab filter
- Harris Lab (3) Apply Harris Lab filter
- Heberlein Lab (9) Apply Heberlein Lab filter
- Hess Lab (3) Apply Hess Lab filter
- Jayaraman Lab (2) Apply Jayaraman Lab filter
- Ji Lab (2) Apply Ji Lab filter
- Kainmueller Lab (2) Apply Kainmueller Lab filter
- Karpova Lab (1) Apply Karpova Lab filter
- Keleman Lab (2) Apply Keleman Lab filter
- Keller Lab (3) Apply Keller Lab filter
- Koay Lab (1) Apply Koay Lab filter
- Lavis Lab (4) Apply Lavis Lab filter
- Lee (Albert) Lab (2) Apply Lee (Albert) Lab filter
- Leonardo Lab (2) Apply Leonardo Lab filter
- Lippincott-Schwartz Lab (12) Apply Lippincott-Schwartz Lab filter
- Looger Lab (13) Apply Looger Lab filter
- Magee Lab (6) Apply Magee Lab filter
- Otopalik Lab (1) Apply Otopalik Lab filter
- Pachitariu Lab (1) Apply Pachitariu Lab filter
- Pastalkova Lab (1) Apply Pastalkova Lab filter
- Pavlopoulos Lab (1) Apply Pavlopoulos Lab filter
- Pedram Lab (1) Apply Pedram Lab filter
- Reiser Lab (1) Apply Reiser Lab filter
- Riddiford Lab (1) Apply Riddiford Lab filter
- Rubin Lab (8) Apply Rubin Lab filter
- Saalfeld Lab (7) Apply Saalfeld Lab filter
- Satou Lab (2) Apply Satou Lab filter
- Scheffer Lab (3) Apply Scheffer Lab filter
- Schreiter Lab (2) Apply Schreiter Lab filter
- Sgro Lab (1) Apply Sgro Lab filter
- Simpson Lab (1) Apply Simpson Lab filter
- Singer Lab (11) Apply Singer Lab filter
- Spruston Lab (4) Apply Spruston Lab filter
- Stern Lab (5) Apply Stern Lab filter
- Sternson Lab (4) Apply Sternson Lab filter
- Svoboda Lab (9) Apply Svoboda Lab filter
- Tervo Lab (1) Apply Tervo Lab filter
- Tjian Lab (1) Apply Tjian Lab filter
- Truman Lab (3) Apply Truman Lab filter
Associated Project Team
Publication Date
- December 2012 (16) Apply December 2012 filter
- November 2012 (16) Apply November 2012 filter
- October 2012 (23) Apply October 2012 filter
- September 2012 (6) Apply September 2012 filter
- August 2012 (13) Apply August 2012 filter
- July 2012 (9) Apply July 2012 filter
- June 2012 (15) Apply June 2012 filter
- May 2012 (13) Apply May 2012 filter
- April 2012 (14) Apply April 2012 filter
- March 2012 (10) Apply March 2012 filter
- February 2012 (19) Apply February 2012 filter
- January 2012 (36) Apply January 2012 filter
- Remove 2012 filter 2012
Type of Publication
190 Publications
Showing 41-50 of 190 resultsA consortium of inhibitory neurons control the firing patterns of pyramidal cells, but their specific roles in the behaving animal are largely unknown. We performed simultaneous physiological recordings and optogenetic silencing of either perisomatic (parvalbumin (PV) expressing) or dendrite-targeting (somatostatin (SOM) expressing) interneurons in hippocampal area CA1 of head-fixed mice actively moving a treadmill belt rich with visual-tactile stimuli. Silencing of either PV or SOM interneurons increased the firing rates of pyramidal cells selectively in their place fields, with PV and SOM interneurons having their largest effect during the rising and decaying parts of the place field, respectively. SOM interneuron silencing powerfully increased burst firing without altering the theta phase of spikes. In contrast, PV interneuron silencing had no effect on burst firing, but instead shifted the spikes’ theta phase toward the trough of theta. These findings indicate that perisomatic and dendritic inhibition have distinct roles in controlling the rate, burst and timing of hippocampal pyramidal cells.
View Publication PageMicroscopic images of specific proteins in their cellular context yield important insights into biological processes and cellular architecture. The advent of superresolution optical microscopy techniques provides the possibility to augment EM with nanometer-resolution fluorescence microscopy to access the precise location of proteins in the context of cellular ultrastructure. Unfortunately, efforts to combine superresolution fluorescence and EM have been stymied by the divergent and incompatible sample preparation protocols of the two methods. Here, we describe a protocol that preserves both the delicate photoactivatable fluorescent protein labels essential for superresolution microscopy and the fine ultrastructural context of EM. This preparation enables direct 3D imaging in 500- to 750-nm sections with interferometric photoactivatable localization microscopy followed by scanning EM images generated by focused ion beam ablation. We use this process to "colorize" detailed EM images of the mitochondrion with the position of labeled proteins. The approach presented here has provided a new level of definition of the in vivo nature of organization of mitochondrial nucleoids, and we expect this straightforward method to be applicable to many other biological questions that can be answered by direct imaging.
We discuss the advantages and challenges of the open-source strategy in biological image analysis and argue that its full impact will not be realized without better support and recognition of software engineers’ contributions to the biological sciences and more support of this development model from funders and institutions.
IL-15 plays a multifaceted role in immune homeostasis, but the unreliability of IL-15 detection has stymied exploration of IL-15 regulation in vivo. To visualize IL-15 expression, we created a transgenic mouse expressing emerald-GFP (EmGFP) under IL-15 promoter control. EmGFP/IL-15 was prevalent in innate cells including dendritic cells (DCs), macrophages, and monocytes. However, DC subsets expressed varying levels of EmGFP/IL-15 with CD8(+) DCs constitutively expressing EmGFP/IL-15 and CD8(-) DCs expressing low EmGFP/IL-15 levels. Virus infection resulted in IL-15 upregulation in both subsets. By crossing the transgenic mice to mice deficient in specific elements of innate signaling, we found a cell-intrinsic dependency of DCs and Ly6C(+) monocytes on IFN-α receptor expression for EmGFP/IL-15 upregulation after vesicular stomatitis virus infection. In contrast, myeloid cells did not require the expression of MyD88 to upregulate EmGFP/IL-15 expression. These findings provide evidence of previously unappreciated regulation of IL-15 expression in myeloid lineages during homeostasis and following infection.
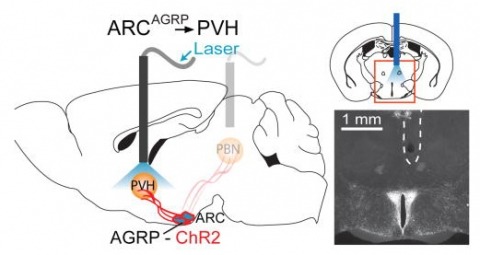
Hunger is a complex behavioural state that elicits intense food seeking and consumption. These behaviours are rapidly recapitulated by activation of starvation-sensitive AGRP neurons, which present an entry point for reverse-engineering neural circuits for hunger. Here we mapped synaptic interactions of AGRP neurons with multiple cell populations in mice and probed the contribution of these distinct circuits to feeding behaviour using optogenetic and pharmacogenetic techniques. An inhibitory circuit with paraventricular hypothalamus (PVH) neurons substantially accounted for acute AGRP neuron-evoked eating, whereas two other prominent circuits were insufficient. Within the PVH, we found that AGRP neurons target and inhibit oxytocin neurons, a small population that is selectively lost in Prader-Willi syndrome, a condition involving insatiable hunger. By developing strategies for evaluating molecularly defined circuits, we show that AGRP neuron suppression of oxytocin neurons is critical for evoked feeding. These experiments reveal a new neural circuit that regulates hunger state and pathways associated with overeating disorders.
The GABA transporters (GAT1, GAT2, GAT3, and BGT1) have mostly been discussed in relation to their potential roles in controlling the action of transmitter GABA in the nervous system. We have generated the first mice lacking the GAT2 (slc6a13) gene. Deletion of GAT2 (both mRNA and protein) neither affected growth, fertility, nor life span under nonchallenging rearing conditions. Immunocytochemistry showed that the GAT2 protein was predominantly expressed in the plasma membranes of periportal hepatocytes and in the basolateral membranes of proximal tubules in the renal cortex. This was validated by processing tissue from wild-type and knockout mice in parallel. Deletion of GAT2 reduced liver taurine levels by 50%, without affecting the expression of the taurine transporter TAUT. These results suggest an important role for GAT2 in taurine uptake from portal blood into liver. In support of this notion, GAT2-transfected HEK293 cells transported [(3)H]taurine. Furthermore, most of the uptake of [(3)H]GABA by cultured rat hepatocytes was due to GAT2, and this uptake was inhibited by taurine. GAT2 was not detected in brain parenchyma proper, excluding a role in GABA inactivation. It was, however, expressed in the leptomeninges and in a subpopulation of brain blood vessels. Deletion of GAT2 increased brain taurine levels by 20%, suggesting a taurine-exporting role for GAT2 in the brain.
Electronic and biological systems both perform complex information processing, but they use very different techniques. Though electronics has the advantage in raw speed, biological systems have the edge in many other areas. They can be produced, and indeed self-reproduce, without expensive and finicky factories. They are tolerant of manufacturing defects, and learn and adapt for better performance. In many cases they can self-repair damage. These advantages suggest that biological systems might be useful in a wide variety of tasks involving information processing. So far, all attempts to use the nervous system of a living organism for information processing have involved selective breeding of existing organisms. This approach, largely independent of the details of internal operation, is used since we do not yet understand how neural systems work, nor exactly how they are constructed. However, as our knowledge increases, the day will come when we can envision useful nervous systems and design them based upon what we want them to do, as opposed to variations on what has been already built. We will then need tools, corresponding to our Electronic Design Automation tools, to help with the design. This paper is concerned with what such tools might look like.
This study provides a new perspective on the long-standing problem of the nature of the decapod crustacean blood-brain interface. Previous studies of crustacean blood-brain interface permeability have relied on invasive histological, immunohistochemical and electrophysiological techniques, indicating a leaky non-selective blood-brain barrier. The present investigation involves the use of magnetic resonance imaging (MRI), a method for non-invasive longitudinal tracking of tracers in real-time. Differential uptake rates of two molecularly distinct MRI contrast agents, namely manganese (Mn(II)) and Magnevist® (Gd-DTPA), were observed and quantified in the crayfish, Cherax destructor. Contrast agents were injected into the pericardium and uptake was observed with longitudinal MRI for approximately 14.5 h. Mn(II) was taken up quickly into neural tissue (within 6.5 min), whereas Gd-DTPA was not taken up into neural tissue and was instead restricted to the intracerebral vasculature or excreted into nearby sinuses. Our results provide evidence for a charge-selective intracerebral blood-brain interface in the crustacean nervous system, a structural characteristic once considered too complex for a lower-order arthropod.
From a genetic screen for Drosophila melanogaster mutants with altered ethanol tolerance, we identified intolerant (intol), a novel allele of discs large 1 (dlg1). Dlg1 encodes Discs Large 1, a MAGUK (Membrane Associated Guanylate Kinase) family member that is the highly conserved homolog of mammalian PSD-95 and SAP97. The intol mutation disrupted specifically the expression of DlgS97, a SAP97 homolog, and one of two major protein isoforms encoded by dlg1 via alternative splicing. Expression of the major isoform, DlgA, a PSD-95 homolog, appeared unaffected. Ethanol tolerance in the intol mutant could be partially restored by transgenic expression of DlgS97, but not DlgA, in specific neurons of the fly's brain. Based on co-immunoprecipitation, DlgS97 forms a complex with N-methyl-D-aspartate (NMDA) receptors, a known target of ethanol. Consistent with these observations, flies expressing reduced levels of the essential NMDA receptor subunit dNR1 also showed reduced ethanol tolerance, as did mutants in the gene calcium/calmodulin-dependent protein kinase (caki), encoding the fly homolog of mammalian CASK, a known binding partner of DlgS97. Lastly, mice in which SAP97, the mammalian homolog of DlgS97, was conditionally deleted in adults failed to develop rapid tolerance to ethanol's sedative/hypnotic effects. We propose that DlgS97/SAP97 plays an important and conserved role in the development of tolerance to ethanol via NMDA receptor-mediated synaptic plasticity.
Learning through trial-and-error interactions allows animals to adapt innate behavioural ‘rules of thumb’ to the local environment, improving their prospects for survival and reproduction. Naive Drosophila melanogaster males, for example, court both virgin and mated females, but learn through experience to selectively suppress futile courtship towards females that have already mated. Here we show that courtship learning reflects an enhanced response to the male pheromone cis-vaccenyl acetate (cVA), which is deposited on females during mating and thus distinguishes mated females from virgins. Dissociation experiments suggest a simple learning rule in which unsuccessful courtship enhances sensitivity to cVA. The learning experience can be mimicked by artificial activation of dopaminergic neurons, and we identify a specific class of dopaminergic neuron that is critical for courtship learning. These neurons provide input to the mushroom body (MB) γ lobe, and the DopR1 dopamine receptor is required in MBγ neurons for both natural and artificial courtship learning. Our work thus reveals critical behavioural, cellular and molecular components of the learning rule by which Drosophila adjusts its innate mating strategy according to experience.