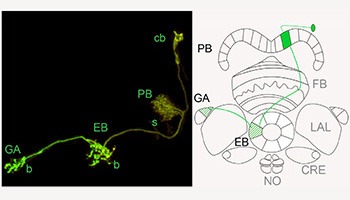
The central complex (CX) comprises four structures (protocerebral bridge, fan-shaped body, ellipsoid body and noduli). The fine structure of the CX was first described in 1989, by U. Hanesch et al. Further anatomy studies have described innervation patterns of single neurons in the central complex (Renn et al. 1999; Young et al. 2009; Lin et al. 2013; Wolff et al. 2014). Nevertheless, the structure and connectivity within the CX remain far from understood.

Three decades of behavioral studies in D. melanogaster, conducted primarily in the laboratories of R. Strauss and M. Heisenberg, reveal a role for the CX in motor control and planning, and the CX has also been implicated in spatial learning and memory (Bouhouche et al. 1993; Seiler et al. 2006; Ofstad et al. 2011) and sleep regulation (Donlea et al. 2011). In other insects, such as the desert locust Schistocerca gregaria, the CX has been shown to play a role in processing polarized light (laboratory of U. Homberg). More recent studies in which the neuronal activity of the CCX have been imaged during behavior, establish a role for the CX in feature detection, orientation tuning and spatial navigation (Seelig and Jayaraman, 2013, 2015).
However, the limited specificity and completeness of tools available for neuroanatomic and behavioral studies has limited our ability to assign observed behavioral phenotypes to specific cell types. In work initiated by Arnim Jenett and continued by Tanya Wolff, we are using MCFO analysis of our GAL4 driver lines to complete the structural analysis of the central complex at the light microscopic and single cell level; in Wolff et al. 2014, we describe the neurons that innervate the protocerebral bridge and we are continuing to analyze the structures of the other portions of the CX in this way. (see gallery for some sample images) Tanya Wolff also plans to work closely with Janelia’s EM efforts over the next few years to generate a CCX connectome.
Using the split-GAL4 intersectional strategy, we are producing a comprehensive set of driver lines with cell-type specific expression (see examples in the section on split-GAL4 lines). These lines can then be used in behavioral experiments, being carried out in several laboratories at Janelia, to significantly increase the precision of structure-function correlations and, consequently, our understanding of the roles of the CX in spatial memory, navigation, sleep regulation and other behaviors. For example, Tilman Triphan in our group (as part of a collaborative effort with Roland Strauss) has shown that inactivation of specific cell types in the protocerebral bridge severely compromises a fly’s ability to orient correctly in a gap crossing task. Working in collaboration with Mike Nitabach’s group at Yale, in a project supported by Janelia’s visiting scientist program, we are studying the role of the CX in sleep.

Specific cell types in the protocerebral bridge are required for proper orientation at a gap. Flies can cross gaps larger than their body length (left). Control genotype flies almost always correctly orient their approach to the gap (center). Flies with certain cell-types in the protocerebral bridge inactivated make frequent errors of orientation (right).