Filter
Associated Lab
- Ahrens Lab (42) Apply Ahrens Lab filter
- Aso Lab (39) Apply Aso Lab filter
- Baker Lab (19) Apply Baker Lab filter
- Betzig Lab (98) Apply Betzig Lab filter
- Beyene Lab (4) Apply Beyene Lab filter
- Bock Lab (14) Apply Bock Lab filter
- Branson Lab (45) Apply Branson Lab filter
- Card Lab (32) Apply Card Lab filter
- Cardona Lab (44) Apply Cardona Lab filter
- Chklovskii Lab (10) Apply Chklovskii Lab filter
- Clapham Lab (10) Apply Clapham Lab filter
- Cui Lab (19) Apply Cui Lab filter
- Darshan Lab (8) Apply Darshan Lab filter
- Dickson Lab (32) Apply Dickson Lab filter
- Druckmann Lab (21) Apply Druckmann Lab filter
- Dudman Lab (34) Apply Dudman Lab filter
- Eddy/Rivas Lab (30) Apply Eddy/Rivas Lab filter
- Egnor Lab (4) Apply Egnor Lab filter
- Espinosa Medina Lab (12) Apply Espinosa Medina Lab filter
- Feliciano Lab (6) Apply Feliciano Lab filter
- Fetter Lab (31) Apply Fetter Lab filter
- Fitzgerald Lab (15) Apply Fitzgerald Lab filter
- Freeman Lab (15) Apply Freeman Lab filter
- Funke Lab (34) Apply Funke Lab filter
- Gonen Lab (59) Apply Gonen Lab filter
- Grigorieff Lab (34) Apply Grigorieff Lab filter
- Harris Lab (48) Apply Harris Lab filter
- Heberlein Lab (13) Apply Heberlein Lab filter
- Hermundstad Lab (17) Apply Hermundstad Lab filter
- Hess Lab (65) Apply Hess Lab filter
- Ilanges Lab (1) Apply Ilanges Lab filter
- Jayaraman Lab (39) Apply Jayaraman Lab filter
- Ji Lab (33) Apply Ji Lab filter
- Johnson Lab (1) Apply Johnson Lab filter
- Karpova Lab (13) Apply Karpova Lab filter
- Keleman Lab (8) Apply Keleman Lab filter
- Keller Lab (60) Apply Keller Lab filter
- Lavis Lab (120) Apply Lavis Lab filter
- Lee (Albert) Lab (29) Apply Lee (Albert) Lab filter
- Leonardo Lab (19) Apply Leonardo Lab filter
- Li Lab (1) Apply Li Lab filter
- Lippincott-Schwartz Lab (85) Apply Lippincott-Schwartz Lab filter
- Liu (Zhe) Lab (51) Apply Liu (Zhe) Lab filter
- Looger Lab (136) Apply Looger Lab filter
- Magee Lab (31) Apply Magee Lab filter
- Menon Lab (12) Apply Menon Lab filter
- Murphy Lab (6) Apply Murphy Lab filter
- O'Shea Lab (3) Apply O'Shea Lab filter
- Otopalik Lab (1) Apply Otopalik Lab filter
- Pachitariu Lab (28) Apply Pachitariu Lab filter
- Pastalkova Lab (5) Apply Pastalkova Lab filter
- Pavlopoulos Lab (7) Apply Pavlopoulos Lab filter
- Pedram Lab (2) Apply Pedram Lab filter
- Podgorski Lab (16) Apply Podgorski Lab filter
- Reiser Lab (43) Apply Reiser Lab filter
- Riddiford Lab (20) Apply Riddiford Lab filter
- Romani Lab (28) Apply Romani Lab filter
- Rubin Lab (101) Apply Rubin Lab filter
- Saalfeld Lab (41) Apply Saalfeld Lab filter
- Satou Lab (1) Apply Satou Lab filter
- Scheffer Lab (36) Apply Scheffer Lab filter
- Schreiter Lab (44) Apply Schreiter Lab filter
- Shroff Lab (21) Apply Shroff Lab filter
- Simpson Lab (18) Apply Simpson Lab filter
- Singer Lab (37) Apply Singer Lab filter
- Spruston Lab (55) Apply Spruston Lab filter
- Stern Lab (68) Apply Stern Lab filter
- Sternson Lab (47) Apply Sternson Lab filter
- Stringer Lab (23) Apply Stringer Lab filter
- Svoboda Lab (131) Apply Svoboda Lab filter
- Tebo Lab (7) Apply Tebo Lab filter
- Tervo Lab (9) Apply Tervo Lab filter
- Tillberg Lab (13) Apply Tillberg Lab filter
- Tjian Lab (17) Apply Tjian Lab filter
- Truman Lab (58) Apply Truman Lab filter
- Turaga Lab (34) Apply Turaga Lab filter
- Turner Lab (24) Apply Turner Lab filter
- Vale Lab (6) Apply Vale Lab filter
- Voigts Lab (1) Apply Voigts Lab filter
- Wang (Meng) Lab (7) Apply Wang (Meng) Lab filter
- Wang (Shaohe) Lab (2) Apply Wang (Shaohe) Lab filter
- Wu Lab (8) Apply Wu Lab filter
- Zlatic Lab (26) Apply Zlatic Lab filter
- Zuker Lab (5) Apply Zuker Lab filter
Associated Project Team
- CellMap (1) Apply CellMap filter
- COSEM (3) Apply COSEM filter
- Fly Descending Interneuron (10) Apply Fly Descending Interneuron filter
- Fly Functional Connectome (14) Apply Fly Functional Connectome filter
- Fly Olympiad (5) Apply Fly Olympiad filter
- FlyEM (50) Apply FlyEM filter
- FlyLight (46) Apply FlyLight filter
- GENIE (40) Apply GENIE filter
- Integrative Imaging (1) Apply Integrative Imaging filter
- Larval Olympiad (2) Apply Larval Olympiad filter
- MouseLight (16) Apply MouseLight filter
- NeuroSeq (1) Apply NeuroSeq filter
- ThalamoSeq (1) Apply ThalamoSeq filter
- Tool Translation Team (T3) (22) Apply Tool Translation Team (T3) filter
- Transcription Imaging (45) Apply Transcription Imaging filter
Associated Support Team
- Anatomy and Histology (18) Apply Anatomy and Histology filter
- Cryo-Electron Microscopy (33) Apply Cryo-Electron Microscopy filter
- Electron Microscopy (11) Apply Electron Microscopy filter
- Fly Facility (39) Apply Fly Facility filter
- Gene Targeting and Transgenics (10) Apply Gene Targeting and Transgenics filter
- Integrative Imaging (10) Apply Integrative Imaging filter
- Janelia Experimental Technology (35) Apply Janelia Experimental Technology filter
- Management Team (1) Apply Management Team filter
- Molecular Genomics (15) Apply Molecular Genomics filter
- Primary & iPS Cell Culture (13) Apply Primary & iPS Cell Culture filter
- Project Technical Resources (32) Apply Project Technical Resources filter
- Quantitative Genomics (18) Apply Quantitative Genomics filter
- Scientific Computing Software (56) Apply Scientific Computing Software filter
- Scientific Computing Systems (6) Apply Scientific Computing Systems filter
- Viral Tools (14) Apply Viral Tools filter
- Vivarium (6) Apply Vivarium filter
Publication Date
- 2024 (102) Apply 2024 filter
- 2023 (177) Apply 2023 filter
- 2022 (166) Apply 2022 filter
- 2021 (174) Apply 2021 filter
- 2020 (178) Apply 2020 filter
- 2019 (177) Apply 2019 filter
- 2018 (206) Apply 2018 filter
- 2017 (186) Apply 2017 filter
- 2016 (191) Apply 2016 filter
- 2015 (195) Apply 2015 filter
- 2014 (190) Apply 2014 filter
- 2013 (136) Apply 2013 filter
- 2012 (112) Apply 2012 filter
- 2011 (98) Apply 2011 filter
- 2010 (61) Apply 2010 filter
- 2009 (56) Apply 2009 filter
- 2008 (40) Apply 2008 filter
- 2007 (21) Apply 2007 filter
- 2006 (3) Apply 2006 filter
2469 Janelia Publications
Showing 2451-2460 of 2469 resultsComparative analysis of multiple genomes in a phylogenetic framework dramatically improves the precision and sensitivity of evolutionary inference, producing more robust results than single-genome analyses can provide. The genomes of 12 Drosophila species, ten of which are presented here for the first time (sechellia, simulans, yakuba, erecta, ananassae, persimilis, willistoni, mojavensis, virilis and grimshawi), illustrate how rates and patterns of sequence divergence across taxa can illuminate evolutionary processes on a genomic scale. These genome sequences augment the formidable genetic tools that have made Drosophila melanogaster a pre-eminent model for animal genetics, and will further catalyse fundamental research on mechanisms of development, cell biology, genetics, disease, neurobiology, behaviour, physiology and evolution. Despite remarkable similarities among these Drosophila species, we identified many putatively non-neutral changes in protein-coding genes, non-coding RNA genes, and cis-regulatory regions. These may prove to underlie differences in the ecology and behaviour of these diverse species.
The hippocampus is essential for episodic memory, which requires single-trial learning. Although long-term potentiation (LTP) of synaptic strength is a candidate mechanism for learning, it is typically induced by using repeated synaptic activation to produce precisely timed, high-frequency, or rhythmic firing. Here we show that hippocampal synapses potentiate robustly in response to strong activation by a single burst. The induction mechanism of this single-burst LTP requires activation of NMDA receptors, L-type voltage-gated calcium channels, and dendritic spikes. Thus, dendritic spikes are a critical trigger for a form of LTP that is consistent with the function of the hippocampus in episodic memory.
Inducible and reversible perturbation of the activity of selected neurons in vivo is critical to understanding the dynamics of brain circuits. Several genetically encoded systems for rapid inducible neuronal silencing have been developed in the past few years offering an arsenal of tools for in vivo experiments. Some systems are based on ion-channels or pumps, others on G protein coupled receptors, and yet others on modified presynaptic proteins. Inducers range from light to small molecules to peptides. This diversity results in differences in the various parameters that may determine the applicability of each tool to a particular biological question. Although further development would be beneficial, the current silencing tool kit already provides the ability to make specific perturbations of circuit function in behaving animals.
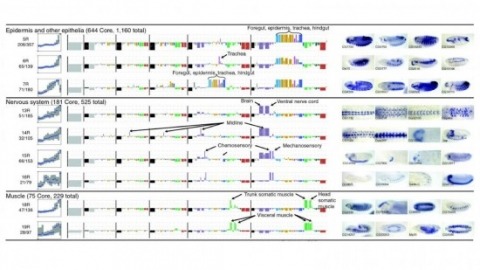
Cell and tissue specific gene expression is a defining feature of embryonic development in multi-cellular organisms. However, the range of gene expression patterns, the extent of the correlation of expression with function, and the classes of genes whose spatial expression are tightly regulated have been unclear due to the lack of an unbiased, genome-wide survey of gene expression patterns.
Staining the mRNA of a gene via in situ hybridization (ISH) during the development of a D. melanogaster embryo delivers the detailed spatio-temporal pattern of expression of the gene. Many biological problems such as the detection of co-expressed genes, co-regulated genes, and transcription factor binding motifs rely heavily on the analyses of these image patterns. The increasing availability of ISH image data motivates the development of automated computational approaches to the analysis of gene expression patterns.
Cortical maps, consisting of orderly arrangements of functional columns, are a hallmark of the organization of the cerebral cortex. However, the microorganization of cortical maps at the level of single neurons is not known, mainly because of the limitations of available mapping techniques. Here, we used bulk loading of Ca(2+) indicators combined with two-photon microscopy to image the activity of multiple single neurons in layer (L) 2/3 of the mouse barrel cortex in vivo. We developed methods that reliably detect single action potentials in approximately half of the imaged neurons in L2/3. This allowed us to measure the spiking probability following whisker deflection and thus map the whisker selectivity for multiple neurons with known spatial relationships. At the level of neuronal populations, the whisker map varied smoothly across the surface of the cortex, within and between the barrels. However, the whisker selectivity of individual neurons recorded simultaneously differed greatly, even for nearest neighbors. Trial-to-trial correlations between pairs of neurons were high over distances spanning multiple cortical columns. Our data suggest that the response properties of individual neurons are shaped by highly specific subcolumnar circuits and the momentary intrinsic state of the neocortex.
This paper presents a new study on a method of designing a multi-class classifier: Data-driven Error Correcting Output Coding (DECOC). DECOC is based on the principle of Error Correcting Output Coding (ECOC), which uses a code matrix to decompose a multi-class problem into multiple binary problems. ECOC for multi-class classification hinges on the design of the code matrix. We propose to explore the distribution of data classes and optimize both the composition and the number of base learners to design an effective and compact code matrix. Two real world applications are studied: (1) the holistic recognition (i.e., recognition without segmentation) of touching handwritten numeral pairs and (2) the classification of cancer tissue types based on microarray gene expression data. The results show that the proposed DECOC is able to deliver competitive accuracy compared with other ECOC methods, using parsimonious base learners than the pairwise coupling (one-vs-one) decomposition scheme. With a rejection scheme defined by a simple robustness measure, high reliabilities of around 98% are achieved in both applications.
In CA1 pyramidal neurons, burst firing is correlated with hippocampally dependent behaviours and modulation of synaptic strength. One of the mechanisms underlying burst firing in these cells is the afterdepolarization (ADP) that follows each action potential. Previous work has shown that the ADP results from the interaction of several depolarizing and hyperpolarizing conductances located in the soma and the dendrites. By using patch-clamp recordings from acute rat hippocampal slices we show that D-type potassium current modulates the size of the ADP and the bursting of CA1 pyramidal neurons. Sensitivity to alpha-dendrotoxin suggests that Kv1-containing potassium channels mediate this current. Dual somato-dendritic recording, outside-out dendritic recordings, and focal application of dendrotoxin together indicate that the channels mediating this current are located in the apical dendrites. Thus, our data present evidence for a dendritic segregation of Kv1-like channels in CA1 pyramidal neurons and identify a novel action for these channels, showing that they inhibit action potential bursting by restricting the size of the ADP.
The functions of cortical areas depend on their inputs and outputs, but the detailed circuits made by long-range projections are unknown. We show that the light-gated channel channelrhodopsin-2 (ChR2) is delivered to axons in pyramidal neurons in vivo. In brain slices from ChR2-expressing mice, photostimulation of ChR2-positive axons can be transduced reliably into single action potentials. Combining photostimulation with whole-cell recordings of synaptic currents makes it possible to map circuits between presynaptic neurons, defined by ChR2 expression, and postsynaptic neurons, defined by targeted patching. We applied this technique, ChR2-assisted circuit mapping (CRACM), to map long-range callosal projections from layer (L) 2/3 of the somatosensory cortex. L2/3 axons connect with neurons in L5, L2/3 and L6, but not L4, in both ipsilateral and contralateral cortex. In both hemispheres the L2/3-to-L5 projection is stronger than the L2/3-to-L2/3 projection. Our results suggest that laminar specificity may be identical for local and long-range cortical projections.
On August 1, 2006 the Howard Hughes Medical Institute's first stand-alone research campus opened at Janelia Farm, near Washington DC. Our mission at Janelia is to do exceptional fundamental research. Our two scientific foci are to understand the function of neural circuits and to develop synergistic imaging technologies. To achieve this we have changed many of the conventions of academic and/or industrial science. The founding director at Janelia is the well-known Drosophilist Gerry Rubin, who has been a central figure in fly molecular, developmental and genomic biology in recent decades. Not coincidentally, we at Janelia fully appreciate the potential of flies to contribute to an understanding of neuronal circuits. Our objectives are ambitious, and in the first ten months of operations at Janelia we have made some good beginnings.